The Masaoki’s Lab
Field Ornithology and Island Biology

The Masaoki’s Lab
Field Ornithology and Island Biology
現在の研究
非学習形質であるフクロウ類の鳴き声の地理的分布は遺伝構造を反映するか?
2014年現在、日本生態学会第61回全国大会 (2014年3月、広島)で講演内容まで進んでいる。
講演要旨 鳥類の分布に与えるケラマギャップの地史的および生態学的時間スケールでの意義
高木昌興(大阪市立大学大学院)、齋藤武馬(山階鳥類研究所)
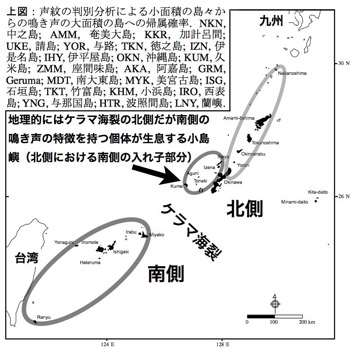
琉球列島のケラマ海裂とトカラ構造海峡で挟まれた島々は中琉球と呼ばれる。更新世前期にはすでに隔離されていたと推察され、非飛翔性の陸生脊椎動物の約80%がこの地域の固有種である。鳥類にもヤンバルクイナなどの固有種が認められ、ケラマ海裂は鳥類の分布境界、蜂須賀線として注目された。しかし在来種に占める固有種の割合は約10%に過ぎず、固有の10亜種も産するが、南・北琉球とそれぞれ約10亜種が共通しており遺伝的交流が推察される。本研究では、琉球列島および南大東島に生息するリュウキュウコノハズクOtus elegansを対象として、鳥類の移動分散の障壁としてのケラマ海裂の持つ意義について検討した。調査は2012-13年に本種の繁殖期である5-7月に行なった。北琉球(中之島)、中琉球(奄美大島、徳之島、伊平屋島、伊是名島、沖縄島、座間味島、久米島)、南琉球(宮古島、石垣島、西表島、与那国島、波照間島)、および南大東島で、捕獲計測・採血、鳴き声の録音を行なった。外部形態、声紋、mtDNA-CO1領域の解析で北・中琉球は区分できなかった。南大東島個体群は完全に独立した。中琉球の大島嶼である奄美大島、徳之島、沖縄島と南琉球の島々の比較では、沖縄島に比較的大きな変異を認めたが、明瞭に区分された。北中と南琉球の遺伝距離は種レベルの違いといえる3%を示した。沖縄島の西側の伊平屋島、伊是名島、座間味島、久米島の個体群の形態と声紋は、中琉球に位置するにも関わらず南琉球の特徴を持っていた。これらの小島嶼の個体群は南琉球のハプロタイプを持っており、その矛盾は遺伝的に裏付けられた。ケラマ海裂は地史的時間スケールでリュウキュウコノハズクの祖先種を分断し、種レベルの変異を蓄積させた。中琉球の小島嶼には生態学的時間スケールで移入が生じていると推察された。
ーーーーーーーーーーーーーーーーー以下、これまでに掲載してきたものーーーーーーーーーーーーーーーーー
鳥のさえずりは生物地理学研究のための形質として利用されている。このような研究は学習によりさえずりを獲得する鳴禽類を材料にしている。具体的には、さえずりを声紋で表現し、シラブルの形状や組み合わせの相違などにより類型化する。それぞれの類型は、学習によって形成されるとの観点から文化的な背景を持つものと考え、方言と表現される。
Marler (2004) は方言を個体群間の遺伝的な相違に帰すことは妥当ではないと示唆した。その後、方言と遺伝構造は対応関係にないことが実際に示された。鳴禽類における遺伝的構造は、文化的な伝播よりも弱いと考えられ、遺伝子流動は文化(方言)と遺伝子進化を乖離させる。一方、Kroodsma et al. (1996) は、鳴き声が学習によって形成されない種、すなわち亜鳴禽類や非スズメ目の鳥類において識別可能な進化的な単位として特に有用であることを示唆している。しかし、依然として実証例はない。
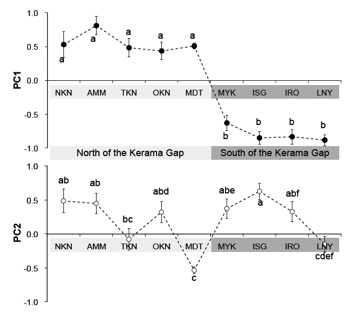
私はリュウキュウコノハズクが生息するほぼすべての島嶼、琉球列島20 島、および南大東島、台湾の蘭嶼島からリュウキュウコノハズクの718 個体の鳴き声を収集し声紋分析を行った(Takagi 2011)。その結果、沖縄島南部のケラマ海峡の南北で声紋に大きな差異を検出した。さらに海洋島に隔離されていると考えらえる南大東島の個体群は明瞭に異なる特徴を獲得していた。この研究は変異がほとんどないと考えられているフクロウ類にも明瞭な地理変異があることを明示した初めての研究であり、この地理変異は氷河性海水準変動による分断の影響が、卓越した移動能力を持つ鳥類にも形質の違いを残す例として貴重な発見となった。
声紋の主成分分析 薄い灰色,ケラマ海裂の北部; 濃い灰色, ケラマ海裂の南部. 凡例は, NKN, 中之島;AMM, 奄美大島;TKN, 徳之島; OKN, 沖縄島; MDT,南大東島; MYK,宮古島; ISG, 石垣島; IRO, 西表島;LNY, 蘭嶼島. Tバー上下の異なるアルファベット間には統計的な有意差が認められた.
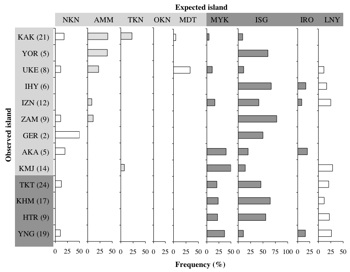
リュウキュウコノハズクの鳴き声には、ケラマ海裂の南北で明瞭な違いがある。そこで遺伝的にもケラマ海裂の南北に違いがあるか確認する。遺伝解析には、ミトコンドリア、核、マイクロサテライトの各DNA を用いる。さらに個体レベルで遺伝的特徴と鳴き声の特徴の関係を明らかにする。ケラマ海裂の北側に位置する沖縄島周辺の小島嶼には、ケラマ海裂の南側にルーツを持つ個体が生息していると予測される。これらの島々の個体群は解析に格好のデータを提供する。
本研究では、発声が学習に因らず生得的な形質であるフクロウ類の一種を用い、鳴き声の地理変異と遺伝構造の対応関係を検討する世界的に類を見ない研究である。これまでの私の研究から明瞭な対応関係が実証される可能性が高いと考えている。実証されれば、世界で初の事例となる。鳴き声と遺伝的帰属の対応関係の実証は、格段に遅れている夜行性フクロウ類の生態学研究に大きく寄与する。
つまり、比較的簡便に利用可能な鳴き声が、個体群構造や移動分散などの研究に有用なツールとして利用することができるようになる。また、ケラマ海裂の北側における鳴き声から見た入れ子構造は、南西諸島の鳥類に特徴な方向性を持った分散による可能性がある。本種の出生地分散は最大でも2km程であり(Takagi unpubl data)、海を越えた能動的な分散は稀である。そこで考えられるのは、台風による受動的な移動である。鳥類では台風による迷行が頻繁に観察される。南西諸島の過去10 年間の台風の経路はケラマ海裂の南側から北の方向にほぼ限定される。近年の細分化傾向にある分類学的観点から、南西諸島における鳥類の分類には混乱が認められている。本研究は、そのような地域における生物・系統地理学研究に新たなに考え方を提起し、他種における遺伝的構造の解析方法に指針を提供する。

