インターネット講座2003 「植物細胞の成長と環境応答」
担当:保尊隆享植物細胞は、葉緑体、液胞、及び細胞壁という特徴的な細胞器官(オルガネラ)を持っています。これらの細胞器官は、植物細胞が分裂・成長し、生命活動を行う上で重要な機能を担っています。植物細胞は、それらを取り巻く様々な環境シグナルに対して敏感に反応し、生命を維持しています。そして、植物細胞の成長調節や環境応答、そして細胞間コミュニケーションには、植物ホルモンが関与しています。以上の過程について解説します。
・序論:細胞とは何だろう
・植物細胞の成長
・植物細胞の環境応答
・細胞機能と植物ホルモン
序論:細胞とは何だろう
はじめに序論では、細胞の基本的な特徴を概括します。そのほとんどは既に皆さんがご存じの事柄ですが、復習を兼ねて一読してください。
1.生物は細胞からなる
すべての生物は細胞から構成される。この考え(細胞説)は、R. HookeやM. Malpighiに始まる多くの形態学的な研究の成果を踏まえて、19世紀前半にM. J. SchleidenとT. Schwannによって確立されました。細胞説は今では至極当たり前の概念ですが、それまで不明確であった単細胞生物と多細胞生物の関係を解き明かし、生物学を自然科学の1分野として発展させる上で大きな意味を持ったといわれています。
2.細胞は外界から独立する
生物が生命活動を行うためには、外界から独立して、外部とは異なる内部環境をつくる必要があります。そのための境界が個々の細胞を取り囲む細胞膜です。細胞膜の主成分は、親水性の基と疎水性の構造とを併せ持つ脂質であり、疎水性部分を内側に集めて両側に親水性部分が位置する脂質二重層と呼ばれる構造をとっています。
内部環境を生命活動に都合のよい状態に保つためには、様々な物質の濃度を外界とは異なったレベルに調節する必要があります。そのために、細胞膜を貫通して存在するタンパク質の通路を利用して物質を輸送します。以前は、小さな分子は脂質二重層を押しのけて通り抜けると考えられていましたが、水ですらタンパク質の穴を通ることがわかってきました。膜を介した物質輸送に関わるタンパク質は、チャンネル、キャリアー(トランスポーター)、そしてポンプに大別されます。チャンネルは、イオンなどの一群の化合物を濃度勾配にしたがって受動的に運ぶ時に働きます。しかし、多くの場合、細胞は必要な物質をもっと蓄積し、不要な(有害な)物質をさらに追い出さなければなりません。このような電気化学ポテンシャルの勾配に逆らった輸送(能動輸送)にはエネルギーを必要とします。ポンプはATP分解などによって高エネルギー状態を作り出して、またキャリアーは、ポンプなどがつくった高エネルギー状態を二次的に利用して、能動輸送を行います。キャリアーは特定の構造の物質を受動的に輸送する際にも働きます。
これらの膜タンパク質でも、脂質と同様に、膜内部に位置する部分は疎水性のアミノ酸によって構成されています。したがって、タンパク質の一次構造(アミノ酸配列)より膜貫通部分の有無及び位置の推定ができます。
3.細胞は分裂して増殖する
生命活動を指令する遺伝子は増殖し、子孫に伝達されますが、遺伝子本体が直接渡されるわけではありません。伝わるのは遺伝子を含む細胞です。「細胞は細胞から」、これは細胞説を構成するもう一つの概念です。
細胞を増やして子孫に伝えるためのしくみが細胞分裂です。細胞分裂のうち核分裂は、体細胞分裂(有糸分裂)と減数分裂とに分けられます。体細胞分裂は、多細胞生物の個体を構成する細胞を増やしたり、無性生殖を行うための細胞分裂で、倍加した染色体が二分することによって母細胞と同様の2n(2組の遺伝子セット、ゲノム)の娘細胞を生じます。一方、減数分裂は生殖細胞(n)を生み出すための細胞分裂であり、有性生殖によって2nに復帰することになります。
細胞分裂は、核分裂に引き続いて起こる細胞質分裂によって完了します。細胞質分裂のしくみは動物細胞と植物細胞とで大きく異なります。
4.細胞は成長する
細胞分裂(核分裂及び細胞質分裂)を終了した細胞(体細胞)は、原形質の容積を増やして成長し、元の大きさを回復します。動物の受精卵の初期発生(卵割)を除いて、ふつう、細胞成長が正常に進まないと次の細胞分裂が起きません。
植物細胞では、細胞器官の一つである液胞に外液を取り込むことによって、細胞質の容積を増やすことなく、数十倍以上の大きさに成長します。進化の中で、独立栄養と同時に固着生活を選択した植物にとって、大きく細胞成長することは非常に重要な意味を持っています。
5.細胞は分化する
受精卵は個体を構成するすべての細胞になる能力を持っていますが、細胞は発生の過程で分化し、専門化して行きます。分化の時期や可逆性は、動物細胞と植物細胞とで大きく異なります。ほとんどの動物細胞は、発生の比較的早い時期に分化を行います。そして、最近注目を集めているES細胞(胚幹細胞:embryonic stem cell)を除いて、一度分化すると他の細胞になることはできません。それに対して、植物細胞の分化は基本的に可逆的であり、容易に脱分化→再分化が可能です。したがって、植物には、あえてES細胞として区別すべき細胞群は存在しません。
6.細胞は機能を担う
細胞は外界から独立して、物質交代やエネルギー代謝を行います。これが正常に進行しないと、生命を維持できません。多細胞生物では、分化に応じて、各細胞が特徴的な代謝系を発達させ、特徴的な機能を担うようになります。
7.細胞器官がさらに機能を分担する
細胞内には、細胞器官(細胞小器官:organelle)と呼ばれる構造体があり、細胞の機能をさらに分担しています。普通は、膜に包まれた細胞内区分を指しますが、機能的には非膜性の細胞器官も重要です。細胞器官の種類や発達の程度には動物細胞と植物細胞とで違いがあり、また単細胞の微生物細胞ではさらに特徴的な分化が見られます。細胞器官の分化の程度は、生物を原核生物(prokaryote)と真核生物(eukaryote)に大別する上でも重要な指標となっています。
一般に、細胞の模式図といえば、細胞という包みの中に様々な種類の細胞器官がぷかぷかと浮いている絵が浮かびます。しかし、実際の細胞器官は細胞骨格によって複雑に連結されると同時に、実にダイナミックに移動、相互変換、並びに相互作用しています。そして、この事が細胞器官の、さらには細胞の機能に大きく影響しています。
8.細胞は環境に反応する
細胞は、外部の環境の変化に反応しながら生命活動を行っています。そのため、外界と接する細胞膜には環境シグナルに対する受容体が存在しています。受容されたシグナルは細胞内に変換、伝達され、細胞の反応を引き起こします。
植物は通常じっとして動かないため、環境に対する反応も鈍いように思われがちです。しかし、固着生活を選択したからこそ、より敏感に環境変化をキャッチし、それに反応するしくみを備えています。そうしなければ、とうに絶滅してしまったことでしょう。
9.細胞は会話する
細胞は細胞膜で包まれて独立していますが、細胞同士は始終連絡を取り合っています。多細胞生物では、その情報を統合することによって、個体全体の生命活動を調節しています。細胞は、連絡手段として、様々な情報伝達物質や生理活性物質を合成し、他の細胞に届けています。また、情報を電流として伝えることもありますが、これはイオンの輸送に他なりません。
多くの植物細胞では、隣接した細胞同士が細胞膜で完全に仕切られておらず、原形質連絡(plasmodesma)と呼ばれる穴が空いています。低分子の物質は原形質連絡を通って移動でき、これが植物細胞間の会話の一法となると考えられています。
10.細胞は進化する
私たちが現在目にする細胞は、40億年に及ぶ進化の歴史を通して形成されてきたものであり、例え簡単な原核細胞といえども、進化を反映した構造と機能を持っています。少なくとも、ミトコンドリアと葉緑体が他の細胞の共生に由来するものであることは確かです。両者は独自の遺伝子とタンパク質合成系を持っています。また、ミトコンドリアは動物細胞にも植物細胞にもありますが、葉緑体は植物にしかありませんから、前者の共生が先行したはずです。最近では、このような共生が進化の過程で何回か起きたことを示す証拠も得られています。
私たち自身の細胞も、40億年の歴史のなれの果てなのですが、これからまだまだ進化して行くのでしょうか。
以上の点を図にまとめて序論を終わりにします。細胞には、今回取り上げなかった特徴もありますから、いろいろと調べてみてください。
植物細胞の成長
成長とは、生物がその種類を問わず、生体成分の量を増し、体を築いてゆく現象です。体のサイズを大きくすることは、他の生物以上に、植物にとって重要です。植物の特徴として、独立栄養と固着生活があげられますが、独立栄養の基盤となる光合成では、葉の受光面積をいかに大きくするかが切実な課題です。また、固着生活においては、周囲の環境から逃げられないため、体を大きくしてその悪影響(他の生物からの被食も含む)を最小限にすることが求められます。植物は、進化の歴史を通して、状況に応じて柔軟かつダイナミックにその体の大きさを変える術を身につけました。以下、そのしくみについて述べます。1.細胞分裂と細胞成長
生物の個体全体の成長は、細胞分裂による細胞数の増加と、細胞成長による個々の細胞の容積増加によってもたらされます。植物の成長の特徴として、両過程が時間的にも空間的にも分離していることがあげられます。植物の体は、軸性といって、上下方向の主軸を中心として形成されますが、その両先端に最も齢が若い細胞が存在しています。そして、先端から基部に向かって齢の勾配ができています。すなわち、先端には細胞分裂部域が、その基部よりには細胞成長部域が、そして最も基部に細胞成熟部域が位置することになります(図1)。また、1つの器官を見ると、細胞分裂はその器官の成長の初期に集中して起こり、成長の後半には主に細胞成長が行われます。
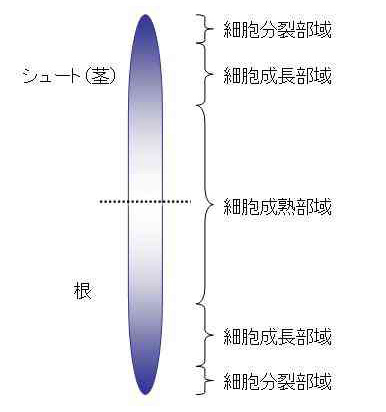
図1 軸に沿った齢の勾配
さて、細胞分裂は、遺伝子を複製しそれを二分する核分裂と、二分した遺伝子セットを別々の細胞に分離する細胞質分裂からなっています。前者の過程は、植物でも、基本的に、動物と同様の道筋に沿って進行します。一方、細胞質分裂の様式に関しては、動物細胞と植物細胞で大きな違いがあります。動物細胞では、分裂赤道面の表層部にアクチンフィラメント(細胞骨格を構成するタンパク質の一種)の働きによってくびれが生じ、それが進行することによって外側から細胞が分けられます。一方、植物細胞では、まず赤道面の中央部に細胞壁(後述)の一種である細胞板が形成され、それが周囲に広がって側壁と結合することによって細胞が分けられます。細胞板形成を担う構造体が隔膜形成体です。隔膜形成体の主な構成要素は、赤道面と垂直に並んだ微小管(他の細胞骨格成分)の束で、これに沿って細胞壁多糖の材料を含んだゴルジ小胞が運ばれ、融合して細胞板を作ります。このようにして形成される細胞板がどの方向にできるかは、植物の形態や分化に大きく影響します。
一般に、植物でも、細胞分裂の頻度は遺伝子のプログラムによって定められており、周囲の環境に応じて植物の成長量が変動する際にもそれほど変化しません。これに対して、細胞成長の程度は大きく変わります。すなわち、植物の個体サイズの柔軟かつダイナミックな変化は、主に個々の細胞のサイズの変化を介してもたらされます。そこで、次に、細胞成長のしくみを見て行くことにします。
2.細胞成長のしくみ
植物細胞の構造上の特徴として、その中央部に液胞と呼ばれる大きな袋(細胞小器官:オルガネラ)を持つことがあげられます。これが、細胞成長量の大きな変化をもたらすしくみとなっています。植物細胞は、しばしば、分裂が終了してから成熟するまでに体積を数十倍にも増やしますが、その間、原形質の量はほとんど増加しません。すなわち、細胞成長は、専ら液胞の体積増加によっていることになります(図2)。そして、液胞が大きくなるのは、細胞外液が液胞内に取り込まれるためだといえます。これを吸水成長と呼びます。
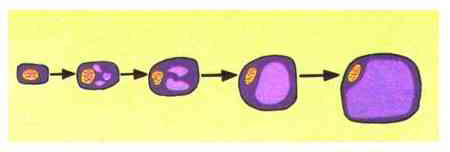
図2 植物細胞の吸水成長
植物の細胞成長の本質が吸水成長であるとしますと、その速度はどのような要因によって調節されるのでしょうか。まず、吸水の原動力ですが、これは液胞液に溶け込んだ溶質によって生み出される浸透圧だと考えられます。浸透圧とは、半透膜を隔てて存在する純水系より水を取り込む力のことです。浸透圧は溶質濃度に比例して大きくなります。一般に、若い植物細胞の液胞液の溶質濃度は、0.4オスモル(理想非電解質溶液と仮定した時のモル濃度)程度ですので、約10バール(=約1メガパスカル)の浸透圧を持つことになります。ふつうの芽ばえでは、浸透圧を生み出す液胞液の溶質(浸透物質)の60%以上がグルコースなどの有機物質です。おもしろいことに、モデル植物として分子生物学で頻用されているシロイヌナズナ芽ばえの浸透物質を測定したところ、有機物質は10%程しかなく、ほとんどがイオン類でした。種子がとても小さくて貯蔵物質の量が少ない植物ですからむべなるかなですが、それにしてもあんな状態でよく生きているものです。
植物細胞は、このように、とても大きな吸水力(=浸透圧)を持っていますが、それを使って自由に吸水するわけには行きません。植物細胞の一番外側には、大きな機械的強度を持つ細胞壁が存在しており、吸水による体積増加を妨げるからです。細胞壁の存在が、植物細胞のもう一つの構造上の特徴です。結局、植物細胞がどの程度吸水できるか、つまり、どのくらい成長できるかは、浸透圧の大きさと細胞壁の伸びやすさ(伸展性)のバランスによって決まることになります(図3)。今までに、様々な条件下で生育している植物で両パラメータが測定されました。その結果、浸透圧は条件にかかわらず比較的安定しており、細胞壁伸展性によって成長速度が規定されるケースが多いことがわかりました。そこで、次に、細胞壁伸展性の調節のしくみを説明します。
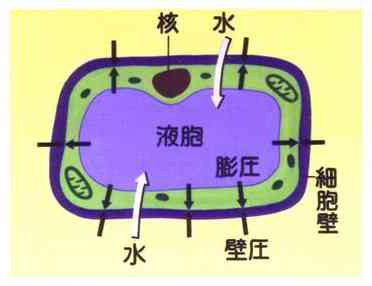
図3 吸水成長の調節機構
ところで、細胞壁の強度によって細胞成長が制限される際、細胞内部には細胞壁強度(圧力)と等価の力が生じます。これを膨圧と呼びます(図3)。一部の教科書には、依然として、膨圧が細胞成長の原動力であり、膨圧が大きくなるほど成長速度も増す、と書かれています。しかし、膨圧は細胞壁が吸水を妨げる程度を反映した力であって、成長速度は、逆に、細胞壁が存在せず膨圧がほとんど0の時に最大になります。したがって、この考えは明らかに誤りです。実際、圧プローブを使って膨圧を測定すると、成長速度が増加する際には、膨圧が一時的に低下することがわかりました。膨圧は、それ自身が積極的に成長調節に関わるというより、浸透圧や細胞壁伸展性など、他の要因のバランスによって受動的に決定されるパラメータであるといえます。
3.細胞成長をもたらす細胞壁変化
さて、ここで、植物細胞壁の構造を説明することが必要になりました。図4は、そのモデルです。細胞壁は、骨格に相当するセルロース繊維とその間に存在するマトリックスの二相構造をとっています。それによって、細胞を守るのに必要な機械的強度と、成長するのに必要な柔軟性の、相矛盾する性質を発揮します。セルロースは、化学的には、β-1,4-グルカンと呼ばれる多糖です。この多糖はお互いに水素結合を形成して会合する性質があり、植物細胞壁中では数十本が集まって束となっています。このセルロースは、地球上に最も多量に存在する有機物質であり(年間生産量1000億トン)、地球温暖化で問題になっている二酸化炭素の最も安定な固定源です。一方、マトリックスは、10種類ほどの多糖及び糖タンパク質の集合体で、セルロース繊維の間を疎らに埋めています。
セルロース繊維は、機械的強度が大きいので、引っ張って伸ばしたり曲げたりすることが困難です。したがって、細胞壁の伸展、すなわち細胞成長は、セルロース繊維と繊維の間を引き離すように起こります。つまり、細胞壁は、図4の左右方向にはほとんど伸展せず、主に上下方向に伸びます。ちょうど、ティッシュペーパーを引っ張った時に起こることと同様です。この事実は、セルロース繊維の配向を変えることによって成長の方向をコントロールできることを意味しています。実際、植物は、そのような方法で成長や形態を制御しています。
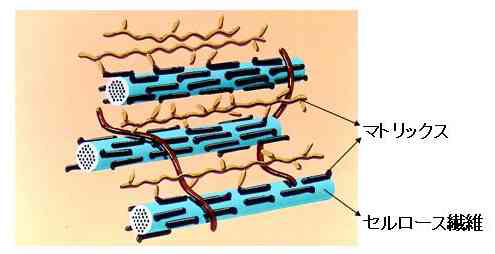
図4 植物細胞壁の構造モデル
セルロース繊維の間を引き離すためには、間を埋めているマトリックス部分に化学的な変化を誘導する必要があります。この過程では、2つの反応が特に重要な働きをしています(図5)。1つは、セルロースとマトリックスの結合を弱めることです。両者の間の結合は水素結合ですので、それを切断すればよいことになります。もう1つは、マトリックス成分間の結合を弱めることです。例えば、多糖鎖を加水分解によって切断して低分子化すれば、マトリックス成分同士の絡み合いが減少し、マトリックス部分全体の粘性が低下して、細胞壁が伸びやすくなります。
このような化学変化は、細胞壁中に存在しているタンパク質(酵素)の働きで起こります。細胞壁の構造は、双子葉植物と単子葉植物(イネ科植物)で異なりますが、双子葉植物では、セルロースとマトリックス間の水素結合の切断にエクスパンシン、マトリックス多糖の低分子化にエンド型キシログルカン加水分解酵素が関わっていることが明らかになりました(図5)。ちなみに、後者の関与は、私たちの研究室の最近の研究でわかったことです。
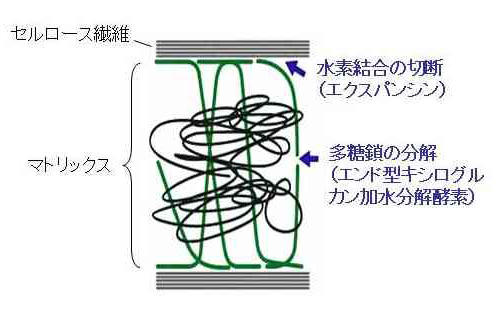
図5 細胞壁の伸展をもたらすしくみ
4.細胞成長の維持
ここまで、植物細胞の成長調節のしくみを説明してきましたが、皆さんの中には、呼吸や光合成、あるいはタンパク質合成のように、生命活動の中心をなす代謝が説明の中に出てこなかったことを意外に思う人がいるかも知れません。以上の話は、あくまで、ある瞬間の成長能力を規定するしくみを解説するためのものです。実際の植物細胞の成長では、もちろん、これらの活性が重要です。例えば、浸透圧の変化によって直接成長速度が変わる例は少ないといいましたが、吸水すると浸透圧がどんどん低下してしまい、すぐにそれ以上成長できなくなりますので、浸透圧を高く維持する機構が必要になります。これを浸透調節といいます。浸透調節において浸透物質を作り続けるためには大きなエネルギーが必要であり、活発な呼吸や光合成によって賄われています。あるいは、細胞壁多糖やその代謝に関与する細胞壁タンパク質の合成や分泌が途絶えると、細胞壁の伸展も止まってしまいます。細胞壁の合成は、成長を長期間維持するために不可欠です。このように、私たちの目に見える実際の植物の成長には、たくさんの過程が複雑に関わっているわけです。
植物細胞の環境応答
今回は環境応答を取り上げます。前回も述べたように、進化の過程で固着生活を選択した植物にとって、周囲の環境の変化を認識しそれに備えることは、自由に移動できる他の生物以上に重要な課題です。そのために、植物は、環境シグナルを敏感に感じ、適切に反応するしくみを獲得してきました。植物の一生に影響する環境要因としては、光、重力、温度、圧力のような物理的な要因、水や各種イオン、化合物のような化学的な要因、そして他の生物による生物的な要因があります。これらの要因は、強さ(高さ、濃度)、方向、あるいは波長(光など)のように、いろいろな属性を持っており、それによって植物の生命活動に影響します。このような環境要因に対して、植物は、まずそのシグナル(信号)を受容(感受)し、それを自分に都合のよい形に変換すると同時に都合のよい場所に伝達し、そして最終的に反応(応答)します。植物細胞の環境応答のしくみを理解するためには、それぞれの環境要因について、各過程を明らかにする必要があります。
今回は、様々な環境要因の中から、重力を取り上げます。重力は、他の要因と違って、地上では大きさも方向も安定しています。そのため、植物は、重力を最も信頼のおけるシグナルとして利用しています。そのような植物の重力に対する応答のしくみを、以下で、曽我康一氏に解説してもらいます。
受講生の皆さん、こんにちは。今回の講義を担当します曽我です。
保尊教授から紹介があったように、今回の講義では植物の重力反応について解説したいと思います。はじめに、なぜ、植物が重力に対して反応しなくてはいけなかったのかについて考えてみましょう。
今から40億年前に海の中で原始的な生命が誕生しました。その後、その生命体は、めざましい進化を遂げましたが、30数億年の間は海から離れることは出来ませんでした。しかし、約5億年前になると大気の上空にオゾン層がつくられ、紫外線などの生物に有害な宇宙線がさえぎられました。そして、水中で生活をしていた植物の一種が太陽の光を多く得ることが出来る陸上に進出しました。陸地の環境は水中と比べるとかなり過酷な環境です。陸に上がった植物は、生存するために様々な困難に立ち向かわなくてはなりませんでした。そのひとつは、浮力のない空気中で、いかに重力に対抗して体を支えるかでした。植物は強固な細胞壁を発達させることによって、この難問を解決しました。この細胞壁は、動物における骨や筋肉の役割をしていると考えられます。このような植物が重力の力に対抗できる体を構築する反応を抗重力反応と呼びます。
陸上では、様々な環境刺激は、絶えずその強さや向きを変えています。例えば、太陽の光は、季節や時間によって、強くなったり弱くなったり、差してくる向きが変わったりします。陸上での固着生活を選んだ植物が、効率的に葉に光を受け、根で水を吸収するためには、自分の姿勢を正しく保つための何らかの情報が必要です。重力は、生命の誕生・進化の過程で、常に一定の方向から一定の大きさで存在していました。このことから、植物は、自分の向きを整えるための基本情報として、重力を選びました。植物は、この最も頼りになるシグナルである重力を感じるための巧みなシステムを作り上げました。このようなシステムをもとに、植物の茎は重力の方向と逆に、根は重力の方向に向かいます。この反応を重力屈性と呼びます。
以上のように、植物は、重力に対抗して体を支える「抗重力反応」と、重力を利用して自らの形を整える「重力屈性反応」の2種類の重力反応を進化の過程で発達させてきました。このように、植物の重力反応には2種類の反応があるのですが、多くの人が注目をしているのは、重力屈性です。おそらく、重力屈性に関しては、高校の生物の教科書にも記載されていると思います。ですから、今回の講義では、はじめに、重力屈性のことを簡単に復習し、その後、もう一つの重力反応である抗重力反応について詳しく説明したいと思います。
重力屈性とは
重力屈性は、植物の茎が重力の反対方向へ、そして根が重力方向に伸長または屈曲する現象です。研究の歴史は非常に長く、1806年にイギリスの植物生理学者のThomas Knightが最初にこの現象について記載して以来、多くの研究者によって研究されてきました。それでは、実際にどのような反応かを見てみましょう。図1にアズキの上胚軸(茎)の重力屈性の様子を示しています。アズキの芽生えを横に倒すと、1時間後には上向きに屈曲を始めているのがわかると思います。時間とともに屈曲と伸長を続け、24時間後には、茎が完全に上を向いているのがわかると思います。本物の植物で重力屈性を見てみたい人は、カイワレダイコンなどを買ってきて、押入などの暗いところに横に倒して置いておくと、観察できると思います。
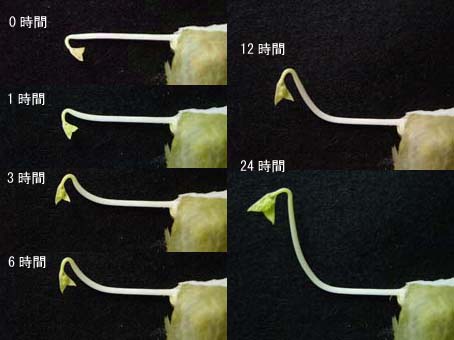
図1 アズキの上胚軸の重力屈性
重力屈性における重力刺激の受容
重力屈性では、重力はどのようにして受容されるのでしょうか。このことを説明するひとつの仮説は約100年前に提唱され、現在でも広く受け入れられています。この仮説は、茎や根を横に倒すと、ある特定の細胞群に含まれる顆粒が重力方向に移動するという観察に基づいています。この顆粒は、デンプンを蓄積したアミロプラスト(デンプン体)と呼ばれる色素体で、ヨウ素ヨウ化カリウム溶液で容易に染色・観察を行うことが出来ます。先程、ある特定の細胞群でと言いましたが、どこで重力を感じているのでしょうか。根では、この点が非常にわかりやすいです。根の先端の0.5 mm程の部分は、根冠と呼ばれ、独特な構造をしています(図2)。根冠は、根の本体にゆるく結合しており、わずかな物理刺激でたやすくはずれてしまいます。根冠がなくても、根は成長し続けます。しかし、根冠を切除すると、横たえてももはや重力方向に曲がることは出来ません。すなわち、根の重力センサーは根冠にあります。ヨウ素ヨウ化カリウム溶液で根を染色すると根冠だけが濃く染まります。身近にある植物の種子を発芽させて、薬局に売っているヨウ素の入ったうがい薬に浸すと、根冠の部分が染まるのを観察できると思います。アミロプラストがどこにあるのかを、さらに詳しく、電子顕微鏡で調べると、根冠の中心付近のコルメラ細胞の中に存在することがわかります(図2)。茎では、内皮細胞(デンプン鞘細胞)にアミロプラストが存在していて、この細胞の中でアミロプラストが重力方向に移動することによって重力刺激が受容されます。このようにして、根や茎で受容された重力刺激は、変換・伝達されて、最終的に屈曲反応を引き起こします。重力刺激の変換・伝達には、植物ホルモンであるオーキシンが関与しています。オーキシンについては、次回の講義で詳しい説明があると思います。重力刺激の変換・伝達の過程に関しては、近年、シロイヌナズナという植物を用いて盛んに研究が行われています。シロイヌナズナを用いた重力屈性の研究に関しては、大学など様々なホームページで公開されていますので、興味のある人は検索してみてください。
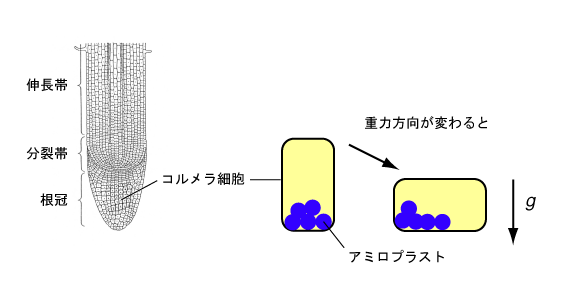
図2 根の構造と重力感受
抗重力反応を研究するには
先程、植物は陸に上がってから強固な細胞壁を持つようになったことを述べました。このことは、藻類と樹木の体を比較すればうかがえることですが、もっと直接的な証拠はないでしょうか。一般的に、生物に対するある要因の影響を調べるときには、その要因の強さ(高さ、濃度)を変化させて、その時の生物の反応を調べます。重力の大きさを変化させるには、どのようにしたら良いのでしょうか。まず、地球上の1 gの重力より小さな重力(微小重力)を作ることを考えてみましょう。落下塔や航空機を用いた自由落下により微小重力環境(いわゆる無重力環境)を作り出すことが出来ます。しかし、微小重力の持続時間は最大でも30秒程度で、植物の成長や細胞壁に対する影響を調べるには短すぎます。それでは、逆に、1 gの重力より大きい重力(過重力)を作ることを考えてみましょう。こちらは、遠心分離機を用いると、容易に、安定した過重力環境を長時間にわたって作り出すことが出来ます。私たちの研究室では、遠心分離機を用いた過重力環境下での植物の成長や細胞壁の変化を解析することによって、植物が重力に対して体を支える仕組みを調べています。また、私たちは、1998年に打ち上げられたスペースシャトルSTS-95において、宇宙の微小重力環境での植物の成長や細胞壁の変化についても研究を行いました。
過重力環境下での植物の成長と細胞壁の変化
地上の1 gよりも大きな重力環境下である過重力環境下では、植物の成長や細胞壁は、どのように変化するのでしょうか。水中から陸上に進出したときに、強固な細胞壁を発達させたように、過重力環境下では、植物は、より強固な細胞壁を発達させる必要があると考えられます。実際、アズキ、シロイヌナズナ、トウモロコシなど、様々な植物を過重力環境下で生育させたところ、重力の大きさが大きくなるにつれて、茎の細胞壁の伸展性が低下しました。すなわち、重力の大きさが大きくなるにつれて、がっちりとした細胞壁を形成しました。前回の保尊教授の講義で説明があったように、細胞壁伸展性が低下すると植物細胞の成長速度は低下します。過重力環境下では、植物は重力に耐えるために細胞壁伸展性を低下させたため、結果として、茎の成長(長さ)は抑制されることがわかりました。さて、今まで、どれぐらいの大きさの過重力環境で植物を生育させたかを話していませんでした。例えば、人間だと、宇宙飛行士などの訓練を受けた人でも、地上の数倍の重力にしか耐えることは出来ません。図3はシロイヌナズナを異なる重力環境下で生育させたときの胚軸の成長を示しています。横軸は、重力の大きさを対数で表しています。右端の点は、何と、300 gです。しかし、植物は重力に強いようで、300 g環境下でも、成長速度は1 gの半分程度に低下しているだけです。もっと驚くことに、エンドウ は10000 g環境下でも、発芽・生育が可能という報告もあります。このような、植物の重力に対する柔軟な適応力が、きっと、水中から陸上への進出を成功させたのでしょう。
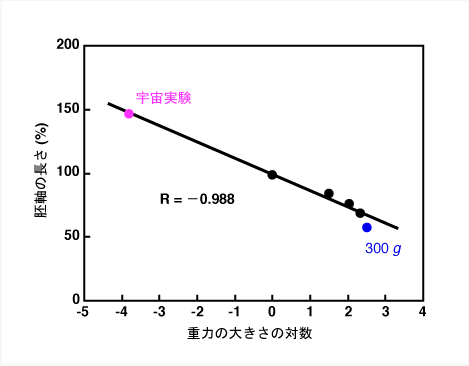
図3 シロイヌナズナ胚軸の成長に対する重力の影響
宇宙の微小重力環境では
過重力環境下では、植物は強固な細胞壁を形成し、重力に対抗しました。宇宙の微小重力環境下では、植物はどのような反応を示すのでしょうか。宇宙では、重力に対抗する必要がないので、柔らかい細胞壁でも体を支えることができると予想できます。このことを確かめるために、私たちは、1998年に打ち上げられたスペースシャトルSTS-95で宇宙実験を行いました(図4)。このSTS-95は、向井千秋宇宙飛行士の2度目の宇宙飛行で、私たちの実験は、向井宇宙飛行士にしていただきました。この実験では、シロイヌナズナとイネを宇宙に打ち上げました。図4Bは宇宙の微小重力環境下でのシロイヌナズナの様子です。ひとつの容器には約1000個体のシロイヌナズナが植わっています。地上で育てたシロイヌナズナは、まっすぐ上を向いていますが(図4A)、宇宙で育てたシロイヌナズナは、いろいろな方向を向いています(図4B)。細胞壁に関しては、私たちの予想通り、シロイヌナズナとイネの細胞壁は、地上に比べて柔らかくなっていました。また、細胞壁が柔らかくなったため、シロイヌナズナとイネの成長は促進されました。図3の左端の点は、シロイヌナズナ胚軸の宇宙での成長を示しています。図3のように植物の成長は重力刺激の対数に応じて変化することがわかりました。細胞壁の固さも成長と同じように重力刺激の対数に応じて変化することが明らかになりました。

図4 宇宙で育てたシロイヌナズナ
どのようにして細胞壁の固さを調節しているのか
過重力環境下で植物は、がっしりとした細胞壁を形成し、宇宙の微小重力環境下では柔らかい細胞壁を形成しますが、次に、この仕組みについて考えてみましょう。ひとつの植物細胞をダンボール箱にたとえて、どうすれば、ダンボール箱が強くなるかを考えてください。一番簡単な方法は、ダンボールを厚くすることです。そうすれば、ダンボール箱は丈夫になります。逆に、薄くすればダンボール箱は弱くなります。実際、植物も、過重力環境下では、厚い細胞壁を作ることで細胞壁を固くしています。逆に、宇宙の微小重力環境下では、薄く柔らかい細胞壁を形成します。ダンボールを固くする、もう一つの方法は、ダンボールの中に針金などを埋め込むという方法です。埋め込む針金の長さが、より長い方が、ダンボール箱は丈夫になります。また、埋め込む針金の量が多いほど、ダンボール箱は丈夫になるでしょう。逆に埋め込む針金の長さを短くし、量を減らすと、ダンボール箱は弱くなります。過重力環境下におかれると、植物は、ある細胞壁多糖の量と分子量(多糖の長さ)を増加させて、細胞壁を固くしています。微小重力環境下では、ある多糖の量と分子量が減少して、細胞壁が柔らかくなりました。双子葉植物と単子葉イネ科の植物では、細胞壁の構造が異なっているので、重力に応答する多糖の種類が異なっています。双子葉植物ではキシログルカン、また、単子葉イネ科植物ではβ-1,3:1,4-グルカンが、その役割を担っています。
抗重力反応における重力刺激の受容
重力屈性では、重力刺激の受容にはアミロプラスト(デンプン体)が関与していることを説明しました。抗重力反応でも、重力刺激の受容にアミロプラストが関与しているのでしょうか。このことを検討するために、アミロプラストが存在しないため重力屈性を示さないシロイヌナズナ突然変異体に対する過重力の影響を調べました。その結果、そのような突然変異体でも、重力屈性を正常に示す野生型と同様に過重力に反応することがわかりました。また、アズキ根では、アミロプラストの存在部位である根冠除去の影響を検討しましたが、根冠を除去して重力屈性能を失わせても抗重力反応は正常に起こりました。しかし、メカノレセプター(機械的刺激受容チャンネル)の阻害剤を植物芽生えに与えたところ、重力屈性は正常に見られましたが、抗重力反応は全く認められませんでした。以上の事実から、重力屈性と抗重力反応における刺激の受容は別々の仕組みによることが明らかになりました。抗重力反応における重力刺激の受容は、アミロプラストを持たない通常の細胞で起こるので、重力刺激は原形質全体の質量として作用し、これが原形質膜上のメカノレセプターによって受容されているのではないかと推察しています(図5)。今後の宇宙実験によって、さらに詳しい情報が得られることでしょう。
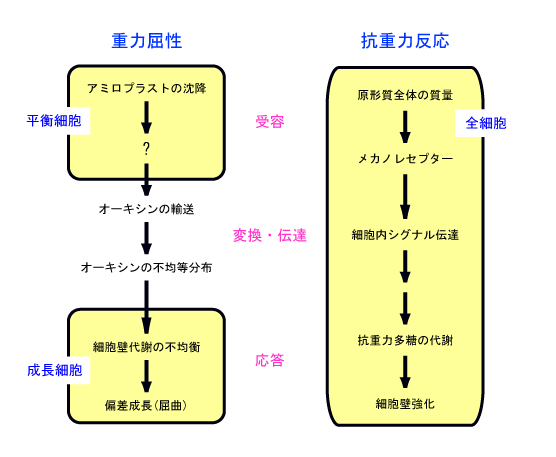
図5 重力屈性と抗重力反応の誘導機構モデル
細胞機能と植物ホルモン
2回にわたって、植物細胞の成長調節と環境応答のしくみを見てきました。今回は、両方の過程で実際に調節の担い手として活躍している植物ホルモンについて解説します。ホルモンという言葉は、もともと「刺激するもの」という意味のギリシャ語に由来し、約100年前から動物学で使われ始めました。動物のホルモンの定義は、「体内の特定の場所で合成され」、「他の場所に運ばれて働く」、「微量で作用を示す物質」、というものでした。1930年代になって、植物の生理活性物質に関してもホルモンという言葉が使われるようになりましたが、植物ホルモンの場合、「特定の場所で合成され」、「他の場所に運ばれて働く」という性質が明確でない物質も存在します。そこで、「植物が自分で合成し、微量で広範な生理作用を示す物質」という、よりゆるやかな定義が認められています。
現在、一般に植物ホルモンとして認められているのは、オーキシン、ジベレリン、サイトカイニン、アブシジン酸、エチレン、ブラシノステロイド、そしてジャスモン酸の7種です。これらの性質について、以下で、若林和幸氏に解説してもらいます。
植物の生活環は、他の生物と同様に、遺伝子の持つプログラムに従って進行しますが、先の講義でも示されたように、その発現が外界の環境によって著しく影響されるという特徴を持っています。植物の生活環の調節において、植物ホルモンは、遺伝子に組み込まれたプログラムが発現される際や、環境刺激(要因)を受容して新たな生理反応(形質)が誘導される時の、エフェクター(作用因子)あるいはメディエーター(介在因子)としての働きを持ちます。植物ホルモンとしては、オーキシン、アブシジン酸、ジベレリン、サイトカイニン、エチレンの5種が古くから認められており、このほかブラシノステロイドとジャスモン酸が、近年、植物ホルモンとして扱われています。7種類という植物ホルモンの数は、植物の生活環の複雑さから考えて必ずしも多いとは言えませんが、実際の植物体では、これらの植物ホルモン間の多様な相互作用により、複雑な生活環の調節がなされていると考えられています。
今回の講義では、上記の植物ホルモンのうち、オーキシン、ジベレリン、サイトカイニン、アブシジン酸、エチレンとブラシノステロイドの6種について解説します。
1:植物ホルモンの発見
[オーキシン]
最も古くから研究されてきた植物ホルモンはオーキシンであり、黎明期の植物ホルモン研究は、そのほとんどがオーキシンに関するものであったと考えられます。オーキシンの発見のきっかけは、植物の屈性の研究でした。1880年にダーウィンは、オートムギの幼葉鞘の先端を覆い光が当たらないようにすると、光方向への屈曲が起こらず、先端部より下の実際に屈曲する部分を覆った場合には屈曲することを報告しました。また、先端部を切り取ると光屈性を示さなくなることから、光の刺激により幼葉鞘の先端部で、屈曲を引き起こす何らかの刺激(シグナル)が誘導されていると考えました。その後、刺激となるものは、ある種の物質であるとする考えがボイセン-イエンセンにより示され、1928年になり、ウエントは刺激を伝える物質を幼葉鞘の先端切片から寒天片に分離しました。1934年に光刺激伝達物質と同様の性質を持つ物質がケーグルらにより人尿から抽出・単離され、その物質がインドール-3-酢酸(IAA)であることが明らかにされました。IAAは、1935年にはカビから単離され、その後、1946年に未熟トウモロコシ種子から単離されて、高等植物自らが生産するホルモンであることが明らかになりました。
[ジベレリン]
オーキシン以外の植物ホルモンに関係する研究は、主に1900年代に入ってから始まりました。次に取り上げるジベレリンは、日本人によって発見されました。イネに発生する病気に馬鹿苗病という病気があり、これに感染すると草丈が徒長し続け穂が出ずに枯れます。1926年に黒沢英一は、馬鹿苗病を引き起こす馬鹿苗病菌の培養液を加熱殺菌して、そのろ過液をイネに与えると馬鹿苗病と同じように徒長することを発見し、馬鹿苗病菌が作り出す物質(毒素)がこの病気の原因であることを明らかにしました。その後、1938年に籔田と住木は、培養液から毒素を単離し、馬鹿苗病菌の学名( Gibberella fujikuroi)からジベレリンと命名しました。しかし、この後、戦争などの影響もあり、ジベレリンの化学構造が決定されたのは1954年になってからでした。この時点では、ジベレリンは菌から取られたものであり、高等植物に存在するかどうかは不明でした。1959年になり、ベニバナインゲンの未熟種子、ゴガツササゲの未熟種子、温州ミカンの徒長枝からジベレリンが単離されて、植物ホルモンの1つであると認められました。現在、100種類以上のジベレリン(類縁化合物)が見つかっています。
[サイトカイニン]
オーキシンやジベレリンが植物細胞の伸長成長を促進するのに対して、細胞の分裂を促進する物質を探す試みが1930年代中頃から盛んに行われました。細胞分裂に関する初期の研究では、ハーバーランドの研究が注目されます。1913年、彼は、ジャガイモの塊茎を切断すると切り口で細胞の分裂が始まり、不定形の細胞の塊(カルス)が形成されることに注目し、傷口で作られる何らかの物質が細胞分裂を引き起こすと考えて、この物質を癒傷ホルモンと名づけました。しかし、彼は、癒傷ホルモンの単離やカルスを人為的に作り出すことは出来ませんでした。1941年になりココナッツの液状胚乳(ココナッツミルク)に、受精直後の胚やニンジンのカルスの増殖を促進する成分が含まれることが示されました。同じ頃、スクーグらは、核酸を構成する塩基の1つであるアデニンが、弱いながらタバコのカルスの細胞分裂を促進する活性を持つことを見出しました。その後、ニシンの精子から調製した古いDNAが強い細胞分裂活性を持つことがわかり、DNAの変性物中に活性物質があることが明らかになりました。1955年になり、この細胞分裂促進物質が単離されて、その化学構造が決定されカイネチンと命名されました。しかし、このカイネチンは、ニシンの精子DNAを人工的に分解してできた化合物であり(天然物ではない)、これが植物体にも存在するかどうかは不明でした。1964年になり、リーサムがトウモロコシの未熟種子から細胞分裂を促進する物質を単離してゼアチンと名づけました。このゼアチンはカイネチンとは異なる物質でしたが、よく似た化学構造を持ち同じ作用を示しました。ゼアチンが単離されたことで、植物体内にも細胞分裂促進物質が存在することが実証されました。その後、よく似た細胞分裂促進物質が様々な植物や微生物から単離され、1965年に、これら一群の細胞分裂促進物質に対し、サイトカイニンという総称が用いられる事になりました。先に挙げたココナッツミルクの活性成分については、1967年にゼアチンとその誘導体であるゼアチンリボシドであることが明らかになりました。ところで、動物の細胞分裂を促進する物質としてサイトカインが知られていますが、これはタンパク質であり、植物のサイトカイニンとは全く違う物質です。
[アブシジン酸]
1930年代になりオーキシン作用の研究が進むにつれて、オーキシンの成長促進作用を抑えるブレーキ役のような物質が植物体に存在することがわかってきました。これがアブシジン酸研究のスタートと考えられますが、その後、アブシジン酸は別々に行われていた成長阻害、器官脱離、休眠芽誘導の3つの生理現象の研究から、それらの原因物質としてほぼ同時期に同定されました。1940年代にヘンベリは、オーキシン作用の阻害物質がジャガイモ塊茎の休眠芽付近に多く含まれ、この阻害物質は塊茎の芽の成長が始まる(休眠が終わる)前に減少することを示しました。1950年代になり、ベネット-クラークとケフォードは、ペーパークロマトグラフィーの手法を用いて植物組織から複数の成長阻害物質を分離し、その1つをインヒビターβと名づけました。後に、ヘンベリの阻害物質もインヒビターβであることがわかりました。しかし、この研究では化学構造の決定には至りませんでした。一方、1960年代のはじめ、ワタ果実の脱離(落果)の研究を行ってきたアディコットらは、器官脱離を引き起こす物質をワタ未熟果実から単離してアブシジンIIと名づけました。このアブシジンIIは、1965年に化学構造が決定されました。同じ頃、ファン・ステベニンクらは、ルーピンマメの果実に花や幼果の脱離を促進する物質が存在することを明らかにしました。もう1つの研究は、同じく1960年代のはじめにウェアリングらによるもので、彼らは、シカモアカエデなどの樹木の休眠芽(冬芽)を誘導する物質の単離を行い、1963年にこの休眠誘導物質をドルミンと名づけました。1967年までに、アブシジンII、ルーピンマメの落果促進物質、ドルミンが同一化合物であることが明らかとなり、この年、アブシジン酸の名称に統一されました。また、インヒビターβについては、アブシジン酸を含む数種類の化合物の複合画分であることが示されました。
[エチレン]
エチレンは、上に示したオーキシン、ジベレリン、サイトカイニン、アブシジン酸とは異なり気体の植物ホルモンです。発見のきっかけは、19世紀の後半頃に照明のために使われていたガス灯の周囲で、街路樹が異常な形態を示すことがたびたび観察されたことです。1901年にネルジュボフは実験室内で育てたエンドウの芽生えが水平方向に伸びることを観察し、この異常は照明用のガス灯からもれたエチレンが原因であることを見出しました。1908年には、カーネーションの花の眠り病(花が閉じて、二度と開かなくなる現象)の原因が、ガス灯からもれたエチレンであることが示されました。同じ頃、エチレンがいろいろな果実の成熟を早める作用を持つことが知られるようになりました。1910年にカズンズによって、オレンジが生成するガスがバナナの成熟を促進する効果を持つことが示され、また、未熟果実と成熟果実を混ぜて貯蔵すると、未熟果実の成熟が早まることが見出されました。1934年になり、ゲインがリンゴ果実からエチレンが発生していることを化学的に証明し、エチレンが植物が生産する気体の生理活性物質であることが明らかになりました。その後、1960年代になり、分析機器のガスクロマトグラフィーが導入されてエチレンの分析(定量)が簡単に行えるようになり、エチレンの生理・生化学的研究が飛躍的に発展しました。
[ブラシノステロイド]
ブラシノステロイドは新しく見つかった植物ホルモンであり、その発見は1970年です。ブラシノステロイドは動物のステロイドホルモンと同じ仲間で、植物で初めてのステロイドタイプのホルモンです。1970年、ミッチェルとマンダヴァは、アブラナの花粉からインゲンマメの第2節間の伸長を促進する物質を見つけ、アブラナ( Brassica napus)の学名から、ブラッシンと名づけました。しかし、含有量が極めて少ないことから、化学構造の決定は出来ませんでした。1978年になり、マンダヴァらが、約230 Kgのアブラナの花粉から15 mgの活性成分を結晶化し、翌年その化学構造が決定されました。その結果、この物質はステロイド骨格を持つことが示されブラシノライドと再命名されました。その後、共通構造としてステロイド骨格を持つよく似た物質が複数単離され、これら化合物はブラシノステロイドという一般名で呼ばれるようになりました。オーキシンやジベレリンなど上に述べた植物ホルモンは、植物が示す様々な生理現象の原因を調べる過程で発見されてきたのに対し、ブラシノステロイド(ブラシノライド)は、ある意味、戦略的に新規の生理活性物質を見つけだすことを目指していた点が、これまでの植物ホルモンの発見とは違っているように思われます。
2:植物ホルモンの構造、生合成と移動
[構造]
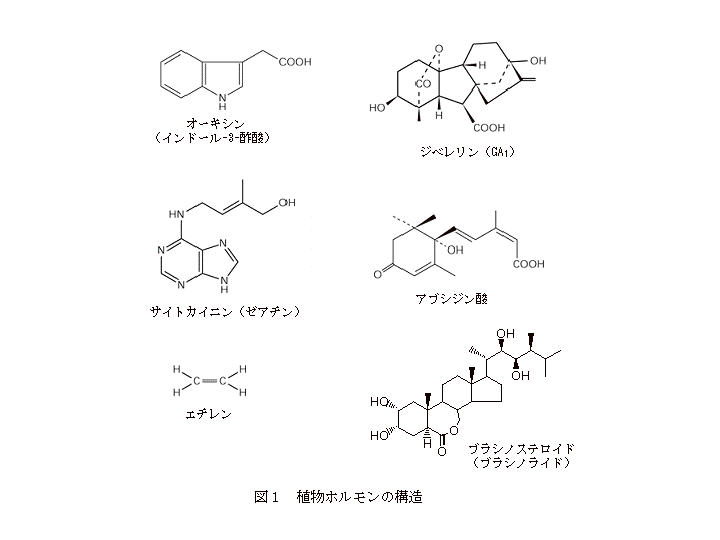
植物ホルモンは基本的に低分子の有機化合物で、その分子量は最も小さいエチレンが28で、最も大きいブラシノステロイド(ブラシノライド)で480程度です。オーキシン、ジベレリン、サイトカイニン、アブシジン酸、エチレンとブラシノステロイドの構造を図1に示しています。
1:オーキシン(auxin)は、ギリシャ語で成長、増加を意味するauxoという言葉が語源になっており、成長を誘導する物質という意味です。オーキシンという名前の化合物は存在せず、オーキシン様作用を持つ物質の総称で、個々の化合物としては、インドール-3-酢酸(IAA、天然物)、2,4-ジクロロフェノキシ酢酸(2,4-D、合成物)、ナフタレン酢酸(NAA、合成物)などが知られています。
2:ジベレリン(gibberellins)は、第1章でも述べたように、イネの徒長成長を引き起こすカビ(馬鹿苗病菌、Gibberella fujikuroi )が産生する物質(毒素)として単離されたことから命名されました。ジベレリンは化合物名であり(gibberellic acid, GA)、現在までに100種類以上のジベレリンが単離されています。それらは全て類似の構造を持つ化合物で、発見順に番号が付けられています( GA1, GA2, GA3, - - - - - )。
3:サイトカイニン(cytokinins)は、cytokinesis(細胞分裂)という言葉が語源になっており、細胞分裂を誘導する物質という意味です。オーキシンの場合と同様に、サイトカイニンという化合物は存在せず、サイトカイニン様作用を持つ物質の総称です。個々の化合物としては、ゼアチン(天然物)、ベンジルアミノプリン(合成物)、カイネチン(合成物)などがあります。
4:アブシジン酸(abscisic acid)は、abscission(脱離、脱落)という言葉が語源になっており、植物の器官脱離を誘導する物質という意味です。アブシジン酸は化合物名であり、数種類の異性体が存在します。
5:エチレン(ethylene)は、化合物(C2H4)名で、ガス状物質です。
6:ブラシノステロイド(brassinosteroids)は、アブラナ(ナタネ)(Brassica napus)の花粉から単離されたことから命名されました。ステロイド骨格を持つ植物で初めてのステロイドホルモンで、類似化合物の総称です。個々の化合物としては、ブラシノライド(brassinolide)などがあります。この他、ジャスモン酸(jasmonates)、ポリアミン(polyamines)も植物ホルモンとして考えられる場合もあります。
[生合成]
動物のホルモンは、体内の特定の場所や細胞で合成されますが、、植物ホルモンの合成は動物の場合とは異なり厳密に決められた合成部位や細胞はありません。しかし、ホルモンの種類により、合成能力の高い部位(組織)は存在します。オーキシンやジベレリンは、頂芽、若い葉、若い茎(茎頂部)、未熟種子で合成能力が高いことが知られています。サイトカイニンは、根の先端部や成長中の芽、未熟種子で合成能力が高く、アブシジン酸は、成熟葉、根、果実、未熟種子などで合成が盛んであると考えられています。エチレンは、成熟果実で生成量が大きいことが知られています。ブラシノステロイドは、花粉、未熟種子、茎の頂芽で主に合成されると考えられています。一般的に、若い時期や組織では、オーキシン、ジベレリンやサイトカイニンの合成が盛んであり、加齢し老化が進むとアブシジン酸やエチレンの合成が大きくなると考えられます。また、環境の変化によっても植物ホルモンの合成量が変わることが知られています。有名な例としては、植物を水分不足の条件に置くとアブシジン酸の合成が増加し、植物の種類にもよりますが、数時間で植物体中の濃度が通常の10倍以上になります。その結果、葉の気孔が閉じて植物体からの水の蒸散が抑えられ水分不足に対応します。
植物ホルモンは、植物体内に比較的ありふれた化合物から合成されます。天然のオーキシンであるIAAとエチレンは、それぞれアミノ酸の1種であるトリプトファンとメチオニンから作られます。ジベレリン、アブシジン酸、ブラシノステロイドとサイトカイニンの側鎖は、イソペンテニル二リン酸を前駆物質として合成されます。また、サイトカイニンのプリン環はATPあるいはADPに由来します。
[移動]
上に述べたように、植物ホルモンでは合成部位と標的部位がはっきりと特定できない場合が多くあります。しかし、茎の伸長部位で作用するオーキシンやジベレリンは頂芽で合成されたものであり、他の植物ホルモンに関しても合成部位とは別の場所に運ばれ作用することがしばしば見られます。サイトカイニンは、根の先端部で合成されて蒸散流によって導管を通り植物体の上部に運ばれると考えられています。植物ホルモンの移動に関して、最も良く研究されているのがオーキシンの輸送です。オーキシンは頂芽で合成されて、根の先端に向けて極性移動することが知られています。このオーキシンの極性移動(輸送)を示したものが図2Aです。放射性のIAAを使った実験から、IAAは茎の上部側から基部側に向かって輸送されますが、その逆の、基部側から上部側へは輸送されないことが古くから知られていました。このオーキシンの極性輸送のメカニズムに関しては、近年、図2Bに示すような細胞膜上にあるオーキシンの取込タンパク質と排出タンパク質が重要な役割を持つことが示されています。茎や根の個々の細胞の上面(上部側)や側面にはオーキシンの取込タンパク質が分布しており、下面(基部側)には排出タンパク質が分布(局在)していることが最近の研究で明らかになってきました。このモデルによると、オーキシンは上面や側面の取込タンパク質によって細胞内に取り込まれ、下面にある排出タンパク質により細胞壁に放出されます。すると、次の細胞の上面や側面の取込タンパク質によって再び細胞内に取り込まれます。このような、取り込みと放出が繰り返されて上部側から基部側に向かっての極性輸送系が形成されると考えられています。また、この取込タンパク質や排出タンパク質は、植物の屈性反応にも深く関わっていることが明らかにされてきています(特に排出タンパク質の局在性が関係しているようです)。
3:植物ホルモンの働き
[生理作用]
イントロダクションでも述べたように、植物ホルモンは、発芽、成長、分化、老化や栄養成長から生殖成長の切換(種子形成)などの植物の生活環の様々なフェーズの調節(制御)に関わっています。また、環境変化に対する植物の応答反応のメディエーターとしての働きを持ち、環境シグナル(例えば光、水、温度など)により、植物体内の植物ホルモン量 が変化して新しい生理形質が誘導されます。植物ホルモンの特徴として、数ナノモルから数マイクロモルという非常に低い濃度で作用する点が挙げられます。また、植物ホルモン作用の濃度依存性は大きく最適型と飽和型の2種類に分かれます。最適型は、作用に最適な濃度があり、それよりも濃度が高くなると作用がかえって小さくなったり、逆に、阻害的に働くタイプのもので、典型的な例がオーキシンです。飽和型は、ある濃度までは作用が大きくなり、その濃度をすぎると作用の大きさがそれ以上に大きくならないタイプで、ジベレリンの作用で見られます。
次に各植物ホルモンの生理作用を示します。
オーキシン:
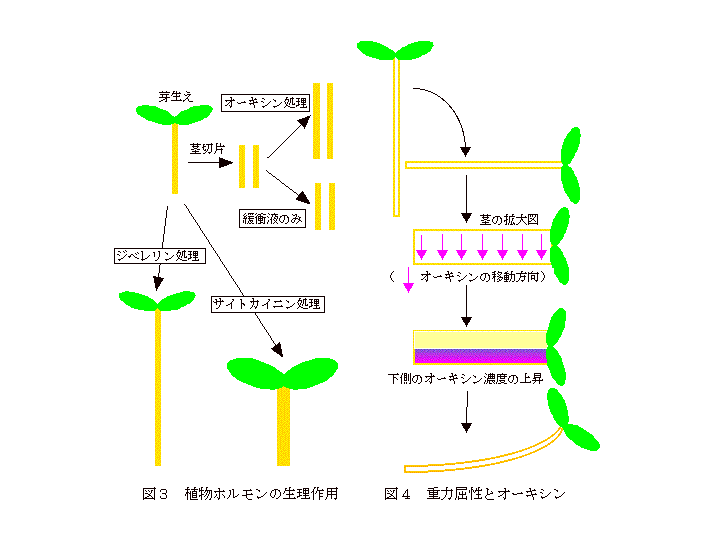
茎伸長促進(主に切片系、図3)、頂芽優勢(側芽成長阻害)、不定根形成誘導、花器官分化、器官脱離抑制、カルス形成誘導(細胞分裂促進:サイトカイニン共存下)
ジベレリン:
茎伸長促進(無傷系、図3)、種子休眠打破・発芽促進、花芽形成誘導、単為結実誘導、果実成長促進
サイトカイニン:
細胞分裂促進(オーキシン共存下)、カルスからのシュート(茎・葉)の誘導、側芽成長促進、茎伸長成長阻害(拡大促進、図3)、葉拡大成長促進、気孔開口促進(蒸散促進)、緑化促進(クロロフィル合成促進)、老化抑制(緑葉の黄化抑制)
アブシジン酸:
種子・芽発芽抑制、休眠(種子・芽)誘導、茎・根成長阻害、器官脱離促進、気孔閉鎖促進、老化促進(緑葉の黄化促進)
エチレン:
茎伸長成長阻害(肥大促進)、器官脱離促進、果実成熟促進、花(花弁)しおれ促進、葉上偏成長誘導、不定根形成促進
ブラシノステロイド:
茎伸長促進、葉身屈曲促進、発芽促進、花粉管伸長促進、ストレス耐性(耐病性、耐塩性、耐冷性等)の促進
[作用機構]
この章では、植物細胞の成長にフォーカスして植物ホルモンの作用機構について述べます。植物の示す生理現象で最も古くから研究されているものが、オーキシン発見のきっかけとなった屈性の研究です。図4には、茎の重力屈性の仕組みを示しています。植物を横たえると茎の中でオーキシンの分布に変化が生じ、茎の下側のオーキシン濃度が高くなります。その結果、茎の下側の成長速度が上側より大きくなり、茎は上方に屈曲して成長すると考えられています。屈性がオーキシンの不均等分布によるという説は、提唱者の名前からコロドニー・ウエント説と呼ばれています。オーキシンの不均等分布のメカニズムについては明らかではありませんが、最近、上の第2章[移動]の項目で述べたオーキシン排出タンパク質の分布変化が関係している可能性が示されています。一方、根では茎とは逆に下方に向かって屈曲します。根の場合もオーキシンの不均等分布が起こり、根の下側のオーキシン濃度が高くなります。しかし、根では成長に必要な最適オーキシン濃度が茎に比べ非常に低いため、下側のオーキシン濃度が最適濃度より高くなり、逆に成長が抑制されて下方に屈曲すると考えられています。
茎の光屈性の場合も同様に、影側でオーキシン濃度が高くなり、影側の成長速度が光側より大きくなることで光側に屈曲すると考えられています。しかし、影側と光側のオーキシン量を調べた結果をみると、オーキシンの不均等分布がみられる場合と、影側、光側でオーキシン量に差が認められない場合とが報告されており、コロドニー・ウエント説だけで光屈性が説明出来るかははっきりとしていません。この光屈性に関しては、光照射により光側で成長阻害物質が合成され、光側の成長速度が低下することで屈曲が起こるという説も提唱されています。
次に、オーキシンが細胞伸長を誘導する仕組みについて説明します。先ず、「植物細胞の成長」で、保尊先生が講義された植物細胞の吸水成長と、細胞壁の構造と役割についての解説を参照して下さい。オーキシンは、細胞膜上にあるプロトンポンプ(プロトン-ATPアーゼ)を活性化し、細胞内から細胞壁への水素イオンの放出を増加させます。その結果、細胞壁内のpHが低下して酸性化します。この時の細胞壁内のpHは4.5~5.5程度になります。細胞壁内には細胞壁の主成分である多糖類を分解する酵素が含まれています。それらの多くは弱酸性(pH 5~5.5付近)に至適pHを持っていることから、細胞壁内が酸性化することでこれら多糖分解酵素の活性が上昇します。これにより、セルロース微繊維の間を埋める多糖類(マトリックス多糖類)の分解が進み多糖類の絡み合いが減少し、マトリックス部分の粘性が低下して、細胞壁全体が緩んで伸びやすくなります。その結果、細胞内への水の取込が増加して細胞の成長がおこります。このようにオーキシンはマトリックス多糖類を分解させ細胞壁を緩ませて細胞伸長を促進することから、細胞壁多糖類の合成が伴わないと細胞壁は薄くなり最終的に伸長は止まります。オーキシンは細胞壁多糖類の分解の促進だけでなく合成も促進することが知られています。これにより、多糖類の分解と合成の繰り返しが維持されて、成長が持続されると考えられています。
ところで、図3に示したようにジベレリンもオーキシンと同じように細胞の伸長促進作用を持っていますが、両者の作用機構はかなり違っています。第5回の説明にあるように、細胞壁中ではセルロース微繊維が縦横に走っており、その構造が維持されています。若い成長中の細胞では、このセルロース微繊維が伸長方向に対して直角(横方向)に並んでいます。細胞が拡大しようとするとき、セルロース微繊維が横方向に「たが」のように並んでいるため横への膨張はほとんどおこりませんが、縦方向へはセルロース微繊維間が緩むことで間隔が広がり伸びることが出来ます。このセルロース微繊維の向き(配向)は、細胞膜の直下(細胞質内)にある細胞表層微小管の向きによって決められていることがわかっています(セルロース微繊維の向きと表層微小管の向きが一致している)。ジベレリンは、この細胞表層微小管の配向を制御して、セルロース微繊維を伸長方向に対して直角に配向させることで伸長成長を促進すると考えられています。さて、ジベレリンの伸長促進の機構にはもう1つあります。それは、細胞の浸透圧を介した機構です。ジベレリンは糖類の転流や取込、デンプンの分解などを促進することで成長部位の細胞の糖濃度を高め、浸透圧を上昇させます。これにより細胞の吸水力が高まり成長が促進されます。この糖濃度の上昇には、ジベレリンによる糖類の加水分解酵素活性の増加が関係していることが示されています。
一方、サイトカイニンは茎などの組織で伸長成長を抑え、拡大を促進します(図3)。これには、上のジベレリンの作用機構で述べた細胞表層微小管とセルロース微繊維の配向が関係しており、これらの配向がジベレリンの場合とは逆になることで拡大が促進されると考えられています。
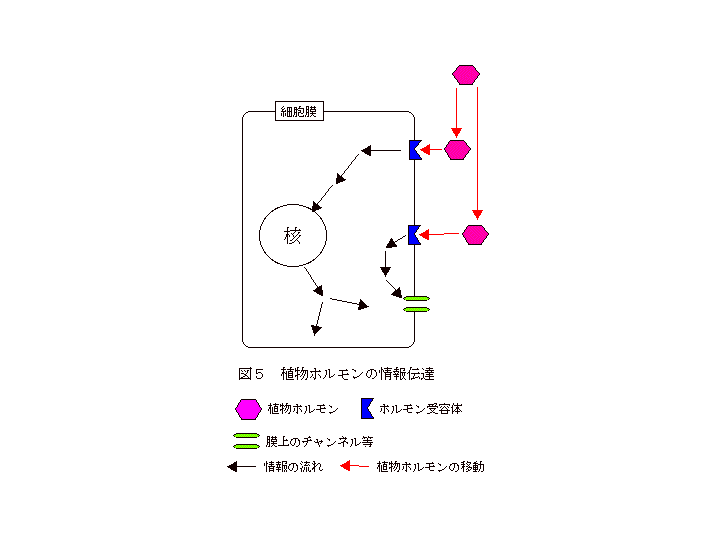
最後に、植物ホルモンの情報伝達系についてふれておきます。細胞膜を透過出来ない高分子の物質と結合させた植物ホルモンを作成して与えた場合でも、ホルモン作用がみられることが知られていました。このことから、植物ホルモンは細胞膜上の受容体(タンパク質)と結合し、その情報が細胞内部へと伝達されることで、機能を発揮すると考えられています。細胞内部へ伝えられた情報は、あるものは核に届き、新たな遺伝子の発現を誘導したり、また、あるものは細胞膜や細胞内膜系の各種のイオンチャンネルなどに作用すると考えられています(図5)。植物ホルモンの受容体に関しては、オーキシン、サイトカイニン、エチレンの受容体が単離されており、サイトカイニンとエチレン受容体については、ホルモンを受容するドメインとその情報を次に伝達するためのドメインを持つことが明らかにされています。今後、他の植物ホルモンのものを含めた多くの受容体が単離され、さらに、受容体の下流に位置するタンパク質が同定されることで、植物ホルモンの情報伝達系の全貌が明らかになると思われます。