研究内容
現在の地球には,名前が付けられた種だけでも約2万種の維管束植物がいる。この多様性は,現在の環境への適応だけでなく,過去の地史学的イベントの結果,成立した。また,過去には体制進化や系統分化の基軸となった分類群が存在したが,そのほとんどは絶滅した。従って,現在の植物多様性を理解するためには,現在と過去をつなぐ研究が欠かせない。そこで,私たちは,古植物学と現生植物学を融合し,維管束植物の体制革新や大系統についての研究を行なっている(山田)。
1.種子の起源と進化
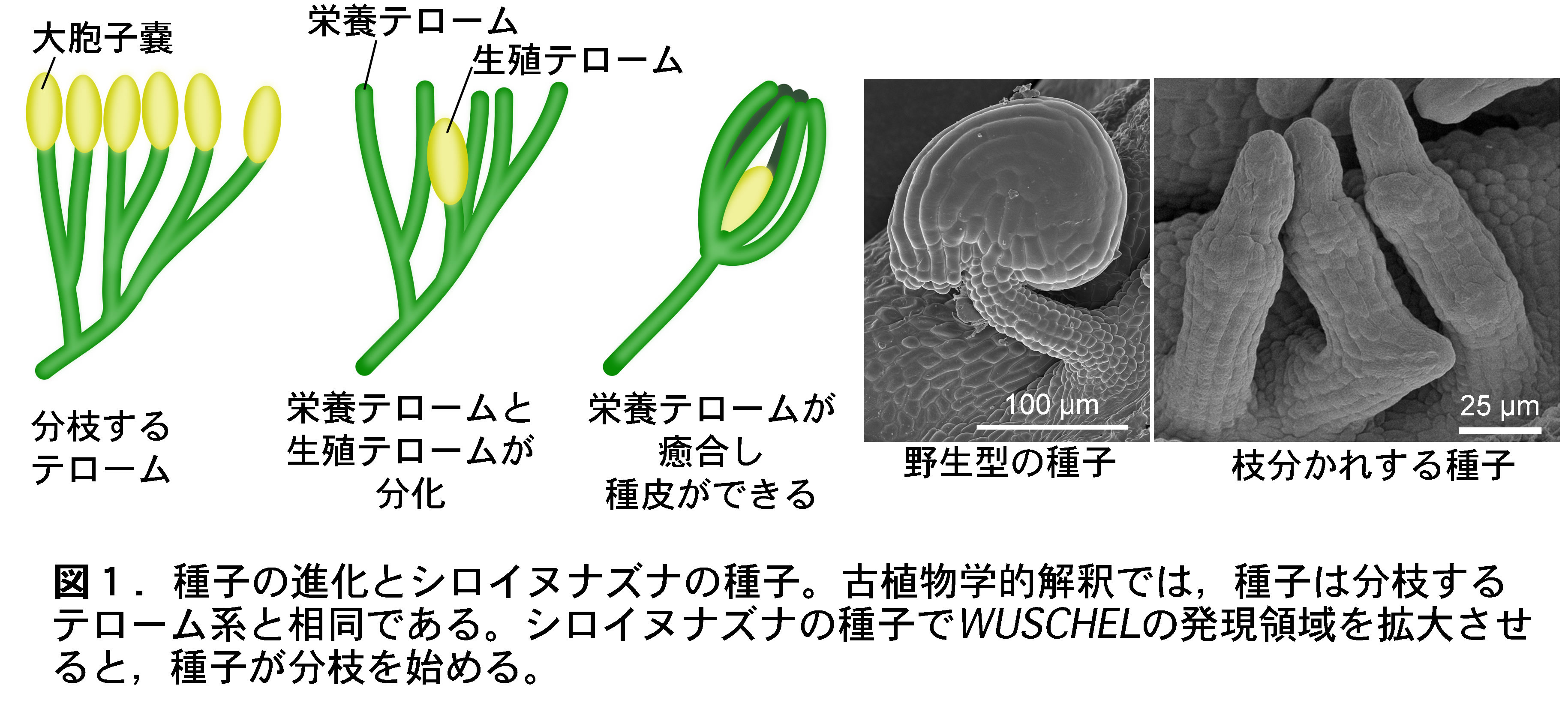
初期維管束植物の体は二叉分岐する軸(テローム)と,胞子嚢だけからできていた。その後,テロームは,栄養テローム(胞子嚢なし)と生殖テロームとに分化した。栄養テロームは癒合して一枚の皮となり,胞子嚢をつける生殖テロームを包んだ。以上が化石から推定される種子の進化である。これに従えば,種子はテローム系そのものと解釈される(図1)。しかし,"分枝する種子"は現存せず,種子とテローム系との相同性を裏付ける発生学的証拠は無かった。
WUSCHELは現生種子植物で同定された軸の先端成長の責任因子である。私たちは,WUSCHELの発現領域が拡大したシロイヌナズナで,種子の軸が分枝することを発見した(図1)。つまり,種子を4億年前のテローム系に先祖返りさせたことになり,両者の相同性を現生種子植物で初めて確認した。
現在,この変異体を使って,4億年前の植物が持った体づくりのメカニズムを解明している。
2.シダの根と種子植物の根の違いを解明する
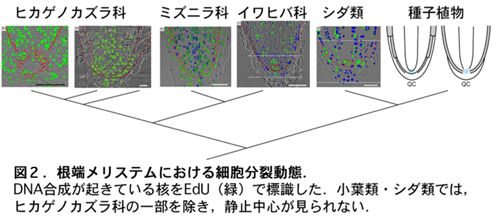
化石記録から,小葉類3科(ヒカゲノカズラ科,ミズニラ科,イワヒバ科),シダ類,種子植物が系統分化した後,各系統で根が平行進化したと推定されている。私たちは,根端メリステムの細胞分裂動態を小葉類3科,シダ類,種子植物で比較し,ミズニラ科,イワヒバ科,シダ類には静止中心(分裂頻度が低くメリステムの維持に働く領域)がないことを示した。一方,ヒカゲノカズラ科の一部には静止中心様の組織があることが分かったが,分裂頻度は種子植物の静止中心よりも高かった(図2)。この結果は,根の多数回起源という古植物学的定説を,根の発生機構の違いから初めて裏付けた。また,この結果は,根の無限成長機構が系統間で異なることを初めて示した。現在は,その違いの原因となる分子メカニズムの解明を行なっている。
3.日本固有の植物相はどのようにできたか?
日本列島は生物多様性ホットスポットの1つで,特に植物の固有性が高い。しかし,第三紀以前に起きたとされる日本の固有植物相の起源は未解明のままである。私たちは,中新世以降の植物化石と現生種との関係解明を進めている。これまでマツ属を中心に研究を進め,中新世最温暖期の日本には,北米やヨーロッパと共通するグループが生育したことが分かった。また最近,中新世最温暖期の水生植物群集を発見したが,その中にはヨーロッパとの共通種が多数含まれるようだ。つまり,最温暖期には全球的に植物の固有性が失われていた。すると,日本の固有植物相を生み出す転機となったのは,中新世最温暖期に続く寒冷化イベントである。そこで,中新世最温暖期前後の植物化石群の変遷を重点的に解析し,日本の固有植物相の成立過程を解明する。
TEL 072-891-2751(山田) FAX 072-891-7199 e-mail tyamada@osaka-cu.ac.jp
研究室所在地 〒576-0004 大阪府交野市私市2000
All Rights Reserved, Copyright © Graduate School of Science / Faculty of Science, Osaka City University