
研究テーマ
2. 刺胞動物の系統と古生物科学
(Phylogeny and Palaeobiology of Cnidaria)
刺胞動物は原始的な後生動物の一群である.中でも「サンゴ類」は,炭酸カルシウム骨格を分泌し,個体性と 群体性の性格をあわせもつ「代表的な造礁性動物」である.サンゴ類は,環境指標に優れ,化石記録も豊富である.現生サンゴを用いた「実験古生物学的な手法 (分子系統学,群体形成様式,生体リズムなど)」も取り入れている.次の観点から研究を行っている.
刺胞動物群の系統発生
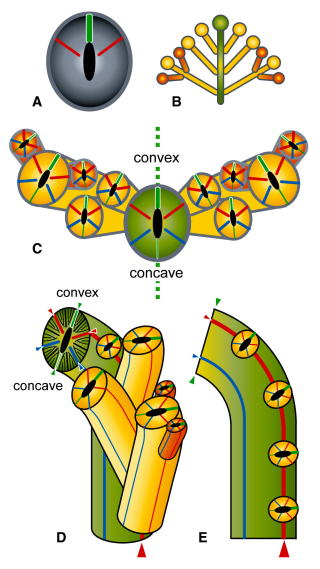
「基本体制の起源や系統発生像」,「四射・床板・六射サンゴの起源と系統関係(カンブリア紀爆発との関係なども)」,「サンゴ類の差別的・選択的な繁栄の歴史」,「六射サンゴの分子系統解析」,「サンゴの記載分類」

イランのジュルファ地域の最上部ペルム系から産する単体四射サンゴPentaphyllum excentricumの連続横断面(Ezaki, 1991).連続的な隔壁の形成で特徴づけられ,六射サンゴへの移行形態は認められない.
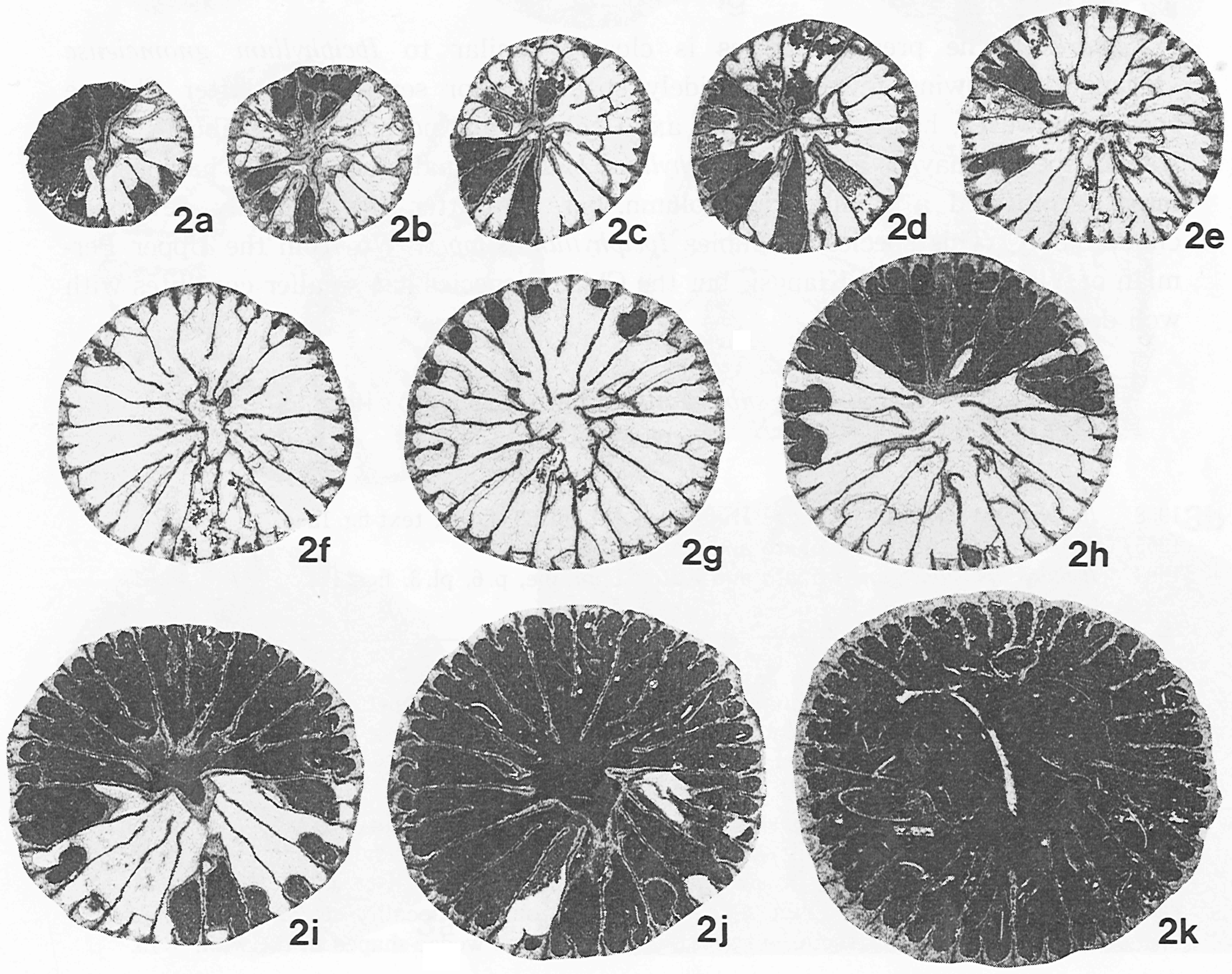
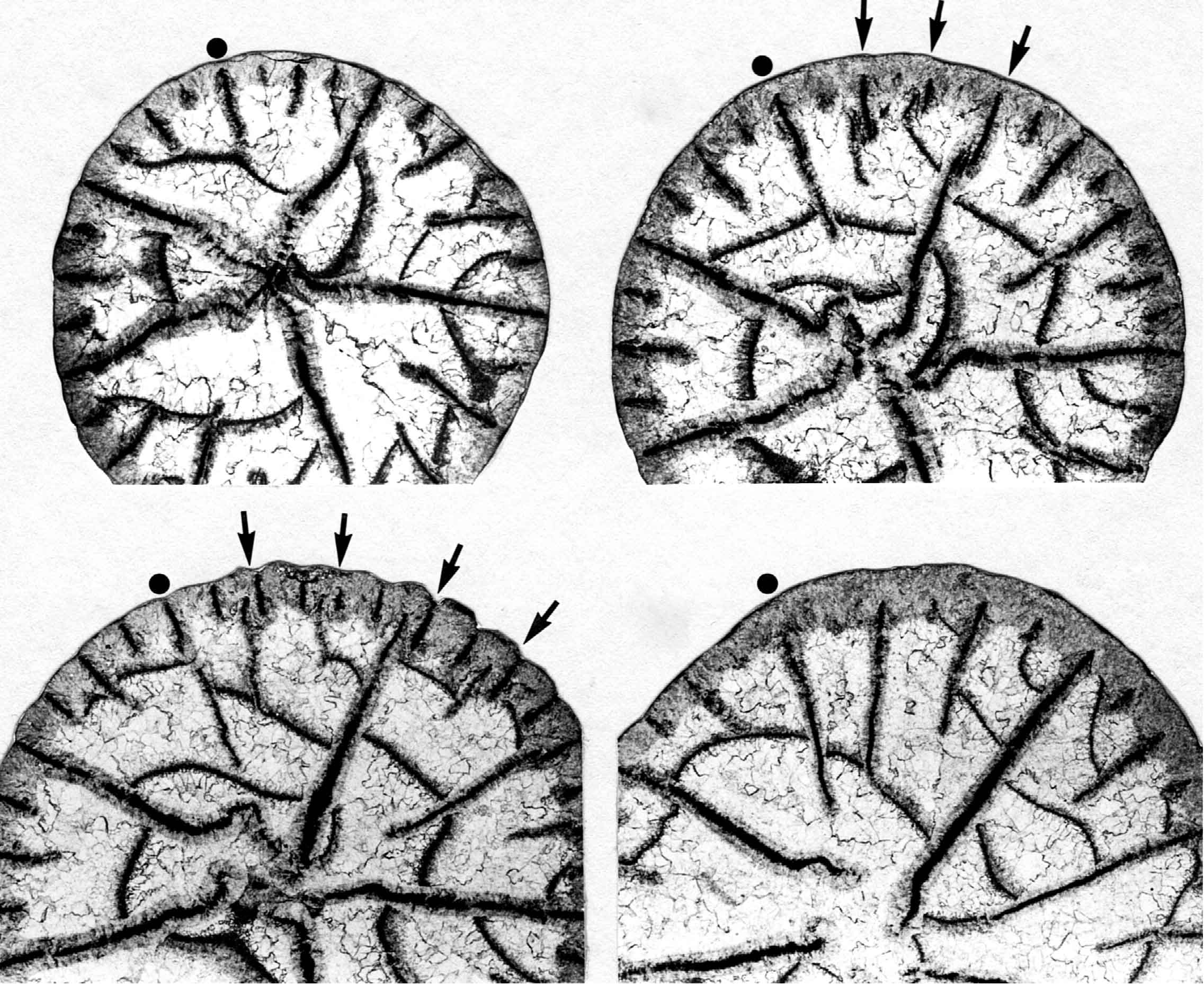
イランのジュルファ地域の最上部ペルム系から産する単体四射サンゴPentaphyllum antractumの連続横断面(Ezaki, 1991).
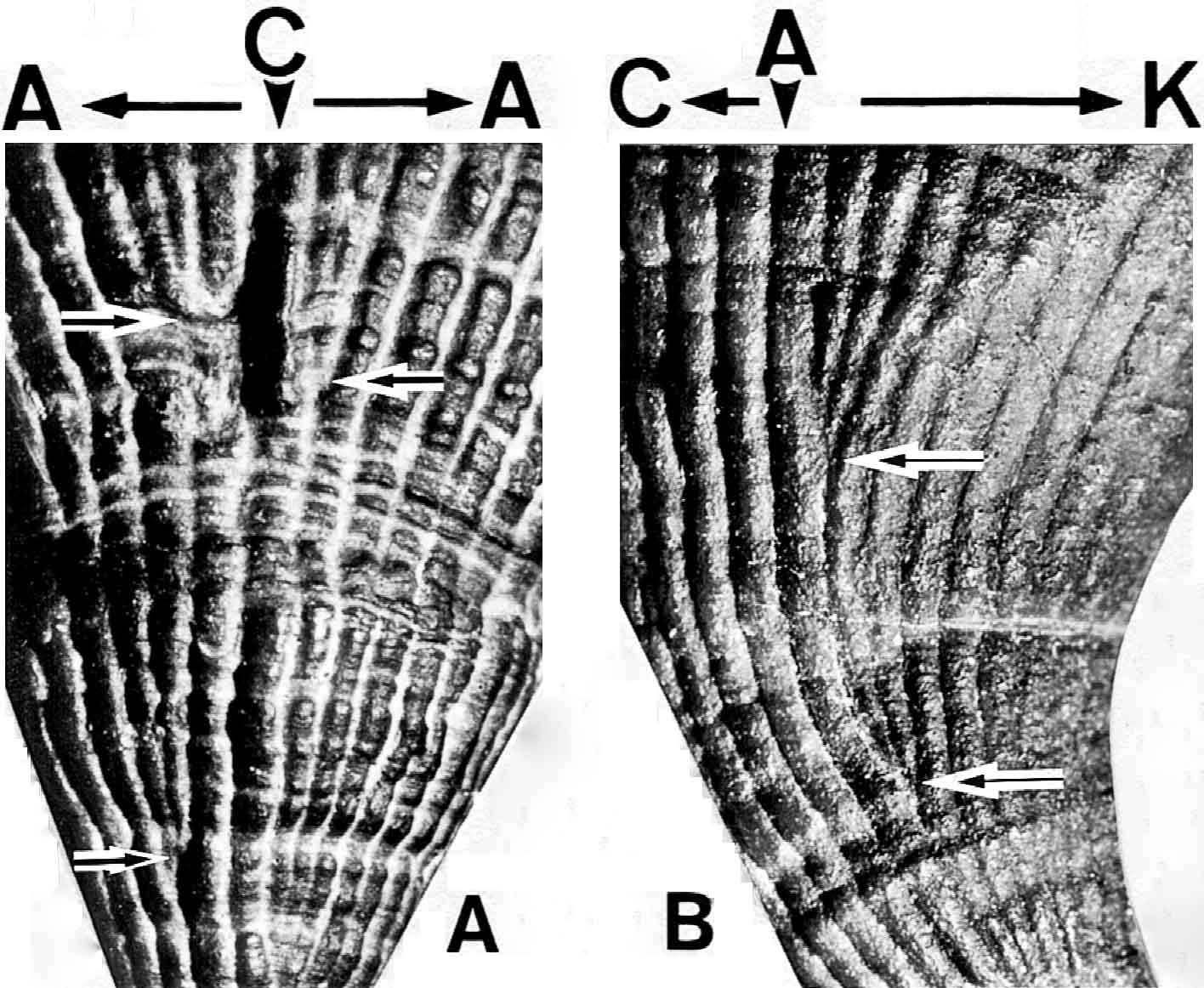
(左)イランのジュルファ地域の最上部ペルム系から産する単体四射サンゴPentaphyllum leptoconicumの壁と隔壁(Ezaki, 1989).成長の途中で一時的に隔壁のずれ(septal dislocation)が生じ(矢印),不規則な隔壁形成が生じているように誤認される.(右)Pentaphyllum leptoconicumの壁表面に形成される隔壁溝.C:主隔壁,A:側隔壁,K:対隔壁(Ezaki, 1989).溝の分岐箇所(矢印)が隔壁の形成箇所(主隔壁の両側と側隔壁の対隔壁側)に相当する.

古生代六射サンゴ:Houchangocyathus wangi.中国貴州省紫雲中部ペルム系(Ezaki, 2000)
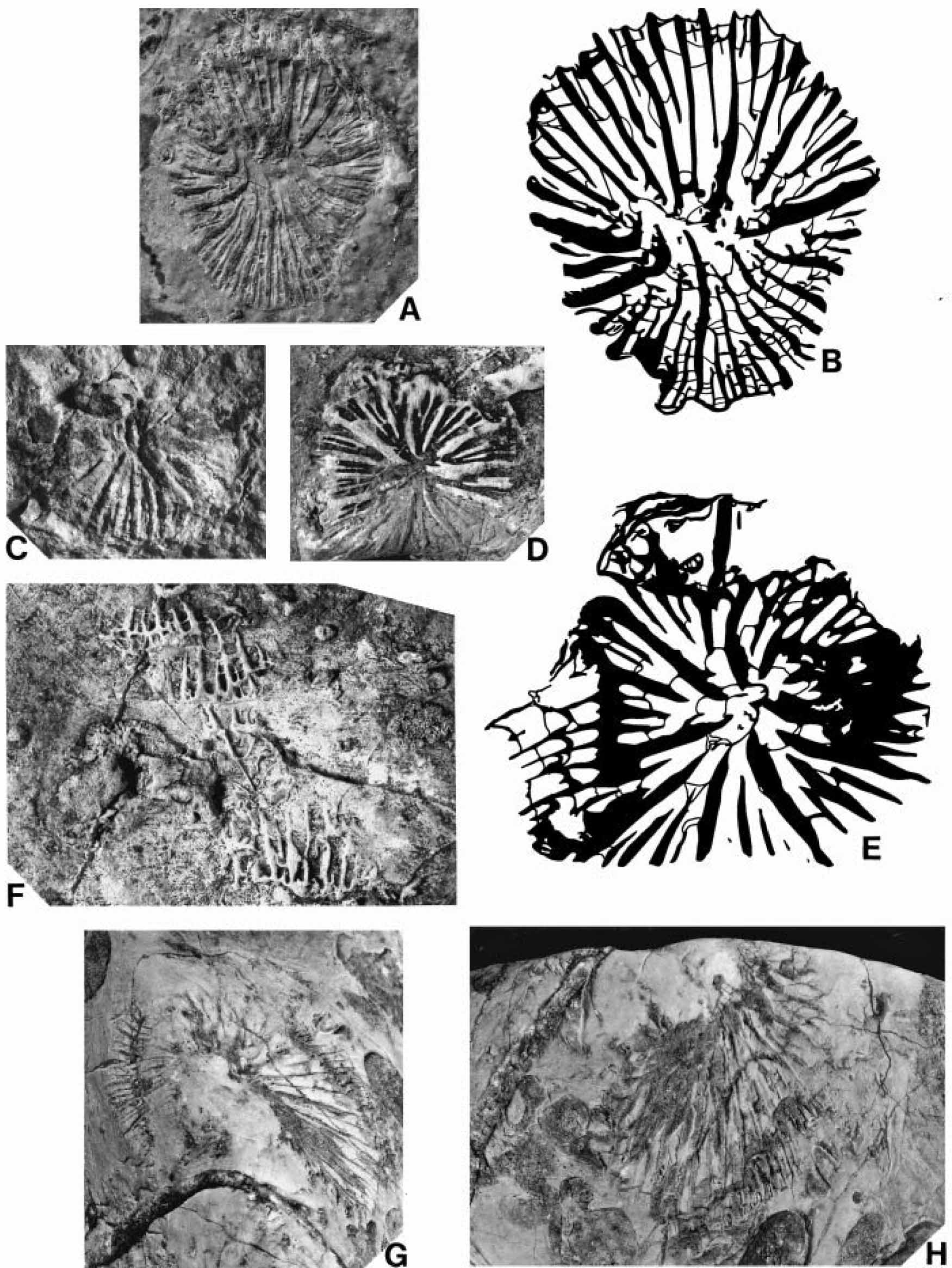
古生代六射サンゴ:Houchangocyathus wangi.中国貴州省紫雲中部ペルム系(Ezaki, 2000)

古生代六射サンゴ:Houchangocyathus yaoi.中国貴州省紫雲中部ペルム系(Ezaki, 2000)
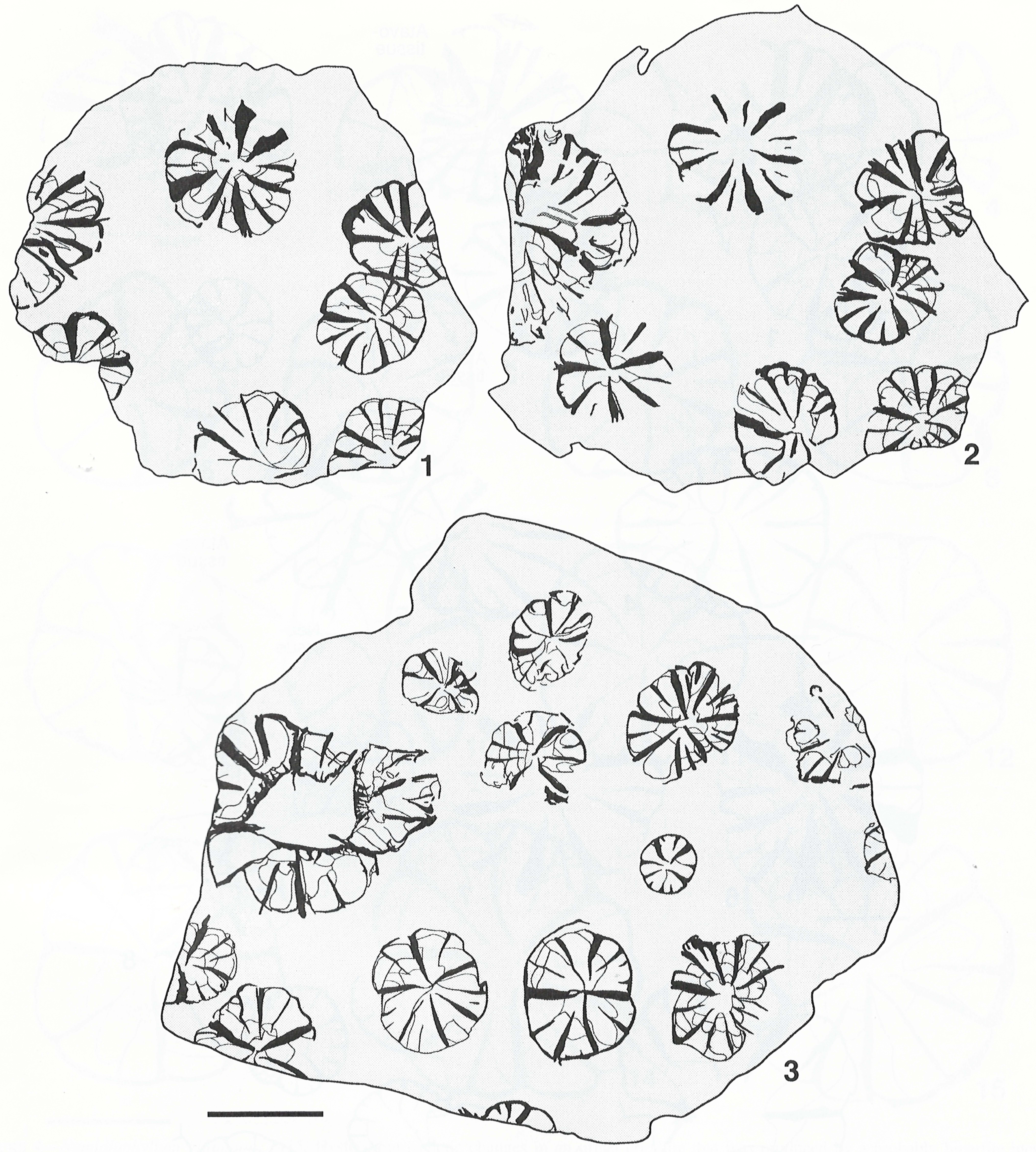
古生代六射サンゴ:Numidiaphyllum gillianum.チュニジア中部ペルム系(Ezaki, 2004)

古生代六射サンゴ:Numidiaphyllum gillianum.チュニジア中部ペルム系(Ezaki, 2004).分裂による無性増殖が顕著である.
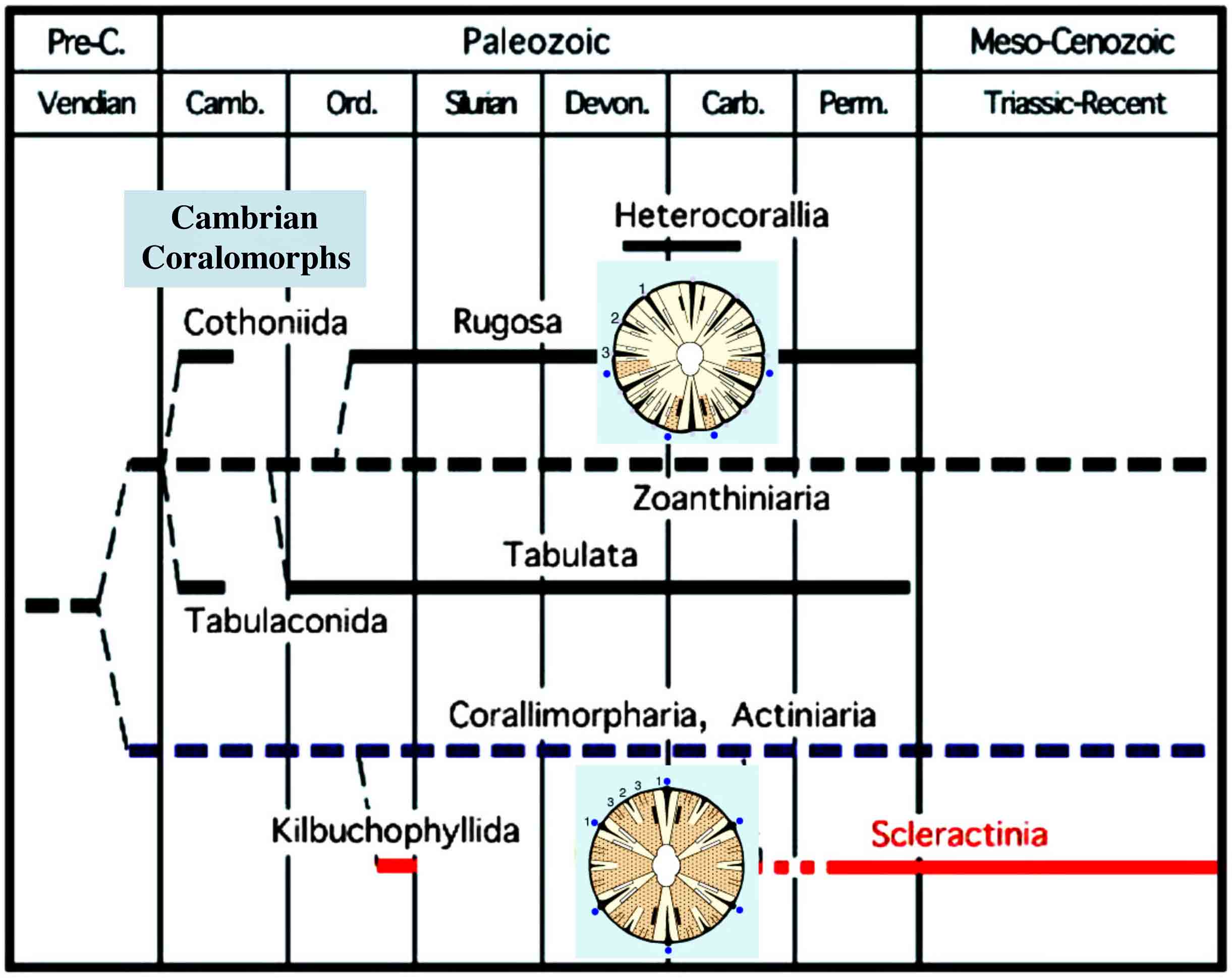
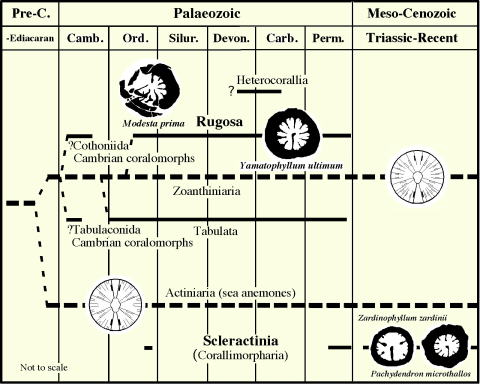
刺胞動物花虫綱の系統関係(Ezaki, 1998).六射サンゴは,古生代に既に出現している!


後期オルドビス紀六射サンゴ(Kilbuchophyllia)を産する地域と地層(スコットランド南部高地 Kirkcolm層)


デボン紀フラニアンのサンゴ化石(ベルギー Vesdre).(左)Lustin層中の四射サンゴ(Disphyllum)群体.(右)Aisemont層中の床版サンゴ(Alveolites)と四射サンゴ(Frechastraea).


(左)Aisemont層(フラニアン)中の群体四射サンゴFrechastraea 生物層の産状:層面(右)群体四射サンゴFrechastraea の産状.(ベルギー アンジ "Tchafornis" park)

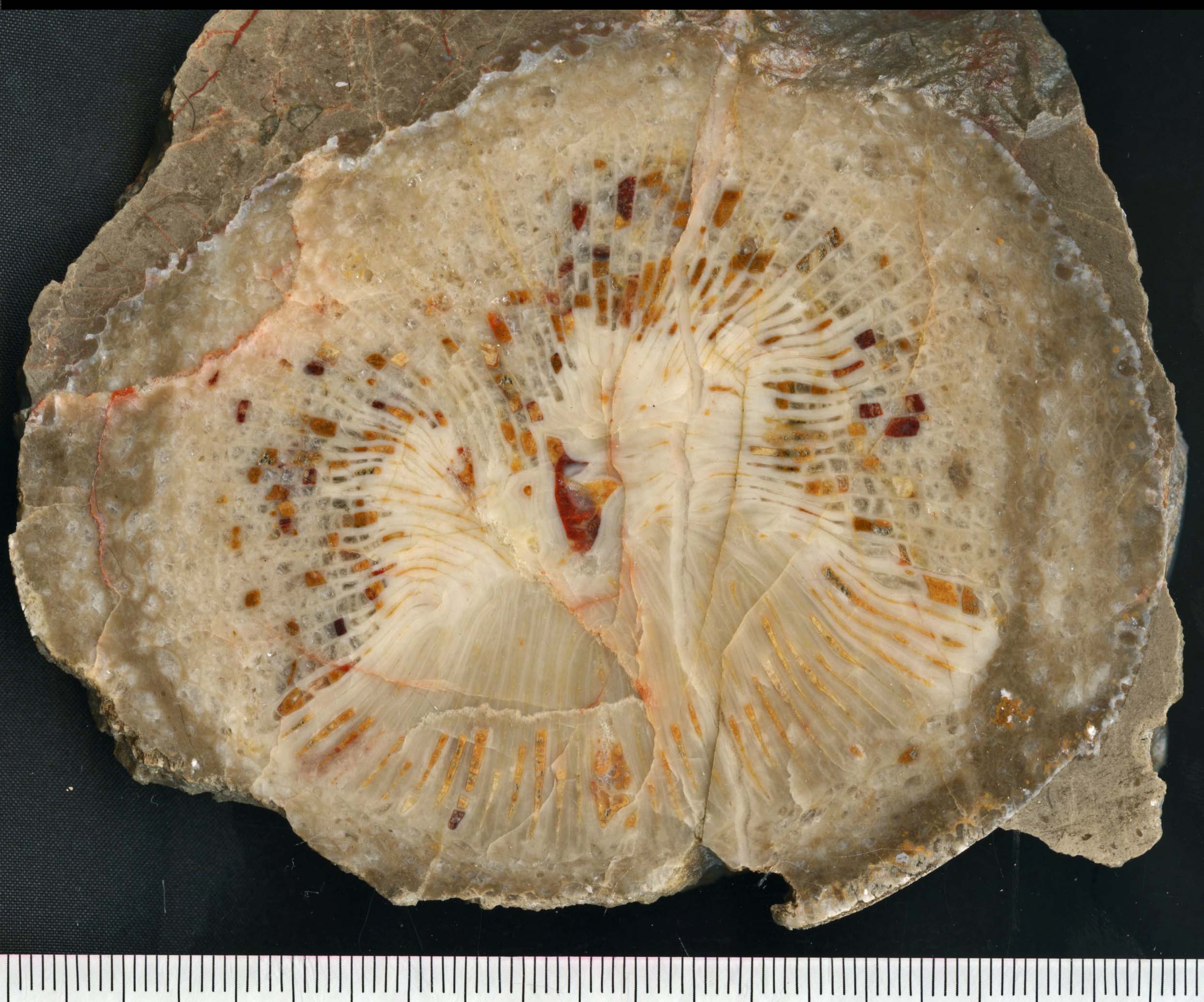
前期石炭紀にテチス域に特徴的に生息したKueichouphyllum(貴州サンゴ).(左)岩手県気仙郡住田町犬頭山産. (右)中国雲南省保山産.


(左)前期石炭紀四射サンゴAphrophyllum grande(Slaughterhouse Creek, Gravesend, NSW, Australia)(右)前期石炭紀四射サンゴSymplectophyllum mutatum(Pinaroo Plain (Caroda), NSW, Australia)
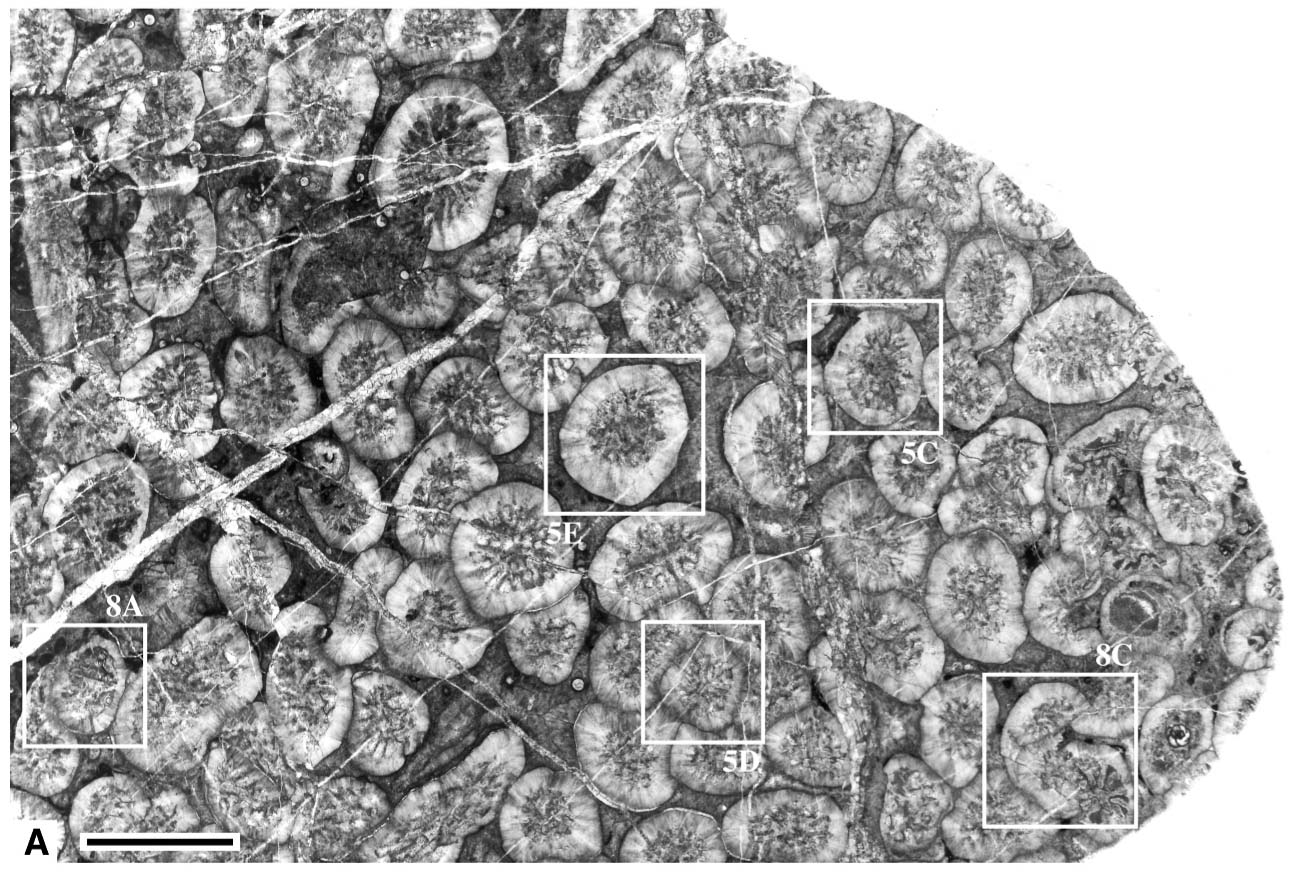
変わった形態組合せをもつ石炭紀群体四射サンゴYamatophyllum ultimum.サンゴ個体はきわめて小さく(平均1.7 mm),2分裂が特徴的である(Ezaki and Kato, 2014).
変わった形態組合せをもつ石炭紀群体四射サンゴYamatophyllum ultimum.サンゴ個体はきわめて小さいが,各形質の発達が特徴的である.白丸は主隔壁(Ezaki and Kato, 2014).
変わった形態組合せをもつ石炭紀四射サンゴと花虫類の系統関係(Ezaki and Kato, 2014).


山口県秋吉石灰岩層群(秋吉帯)から産する四射サンゴ.(左)Nagatophyllum satoi, Carcinophyllum enome. Nagatophyllum satoi帯.(右)Pseudopavona sp. Millerella yowarensis帯〜Pseudostaffella antiqua帯.


山口県秋吉石灰岩層群(秋吉帯)から産する四射サンゴ.(左)Amygdalophylloides sp. Pseudostaffella antigua帯.(右)Akagophyllum sp. Pseudofusulina ambigua帯.
岡山県日南石灰岩(秋吉帯)から産するPseudopavonidae科四射サンゴ.(左)Hiroshimaphyllum sp.(右)Ozakiphyllum sp.

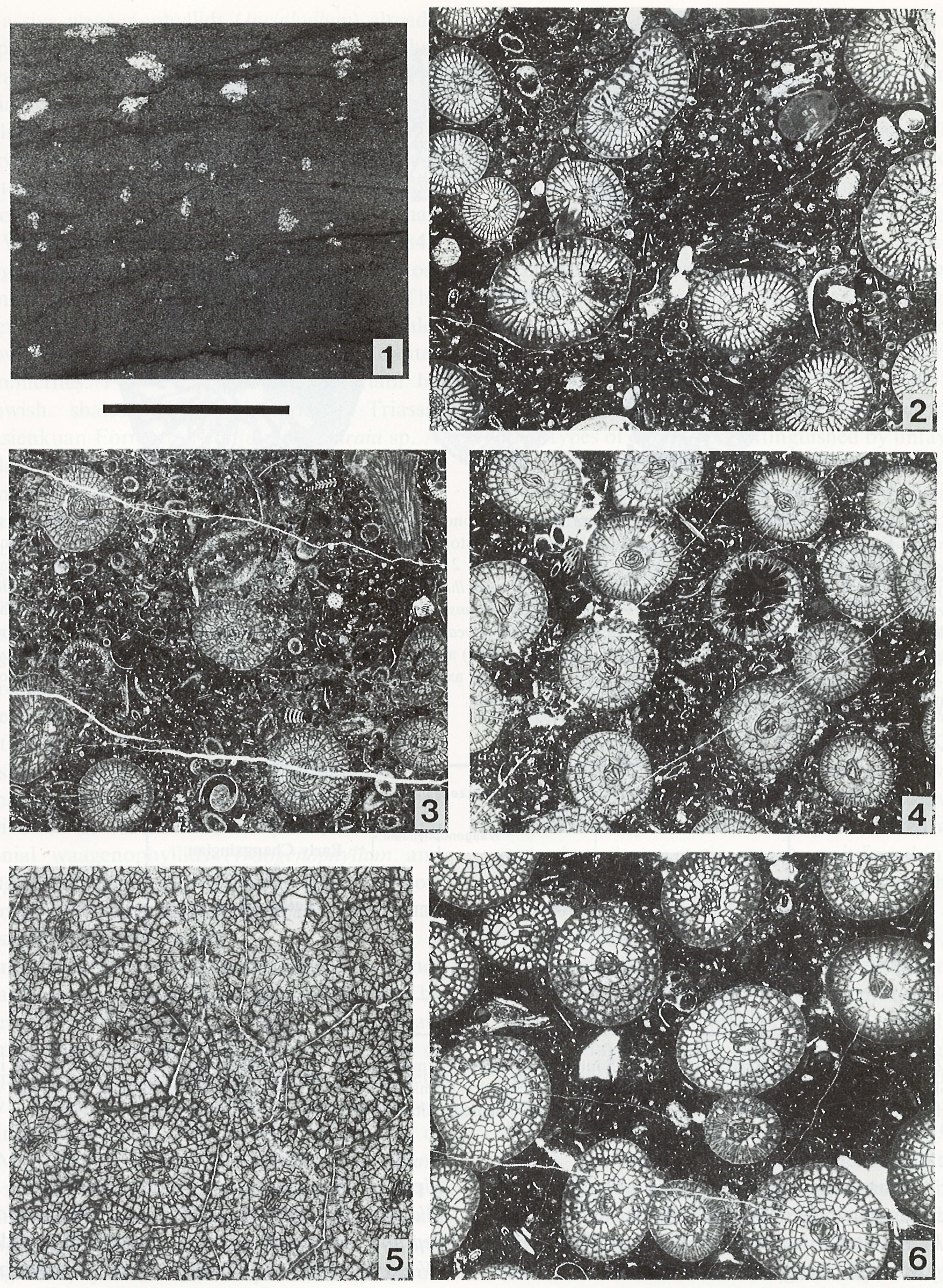
岡山県日南石灰岩(秋吉帯)から産するPseudopavonidae科四射サンゴ.(左)Ibukiphyllum densum.(右)Ibukiphyllum sp.

(左)Pseudopavonidae科四射サンゴPseudopavona taisyakuana. 岡山県日南石灰岩(秋吉帯).(右)Pseudopavonidae科四射サンゴは日本列島の特定の地塊に固有に産する(Kato and Minato, 1975).石炭紀当時の古太平洋の古地理や古海況を考える際に重要な情報を提供する(Ezaki et al., 2007).


前期ペルム系塊状四射サンゴwaagenophyllids(Tabas, Iran)

イランの中部ペルム系から産出する群体四射サンゴ:Lonsdaleoastraea iranica(Ezaki, 1991)
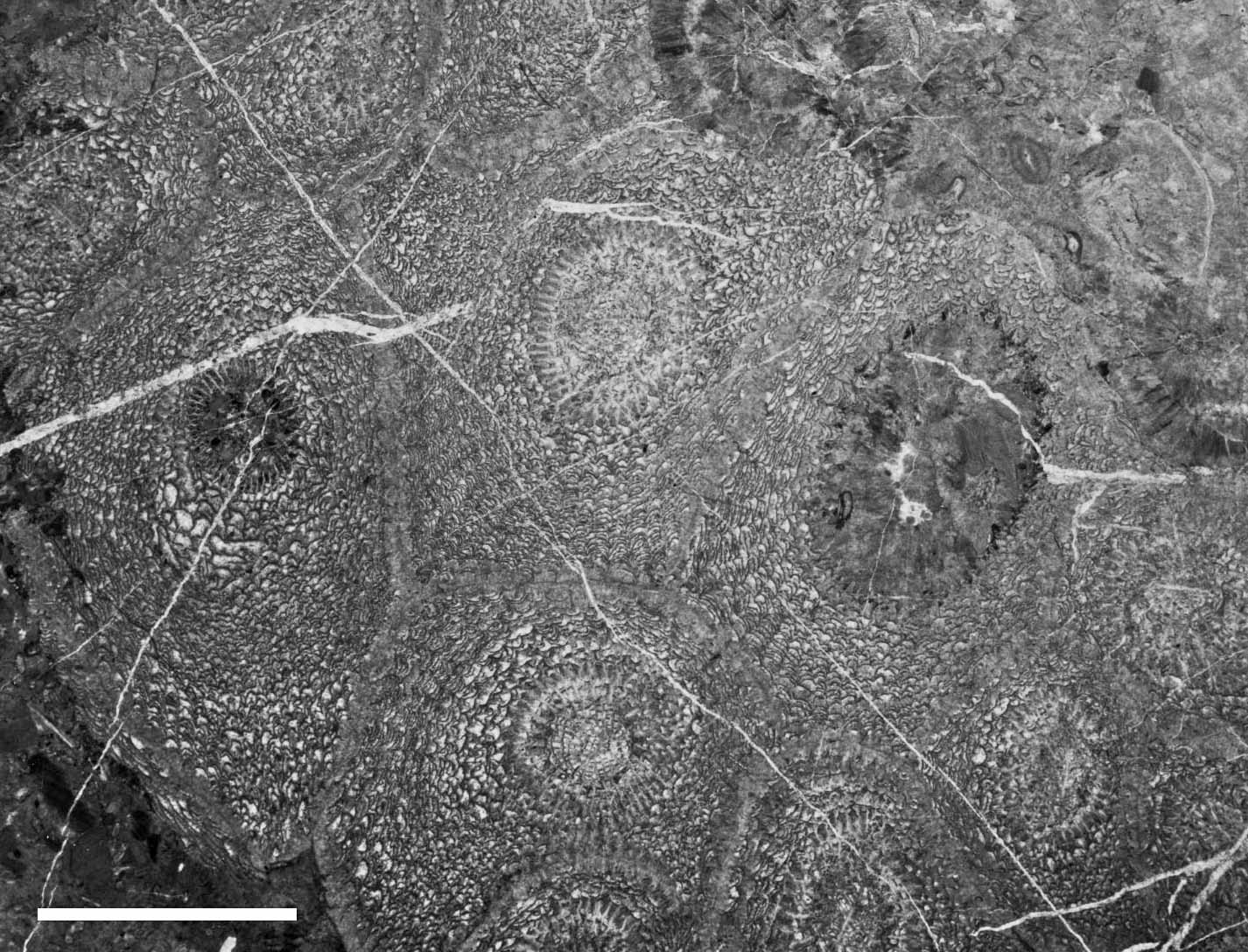
南中国の上部ペルム系の岩相と産出する四射サンゴ(Ezaki, 1994)

南中国における絶滅前の四射サンゴの産出状況.単体と枝状四射サンゴは最上部ペルム系まで産する(Ezaki, 1994).煤山(Meishan)セクション以外では,ペルム系とトリアス系の接触関係は不整合である.
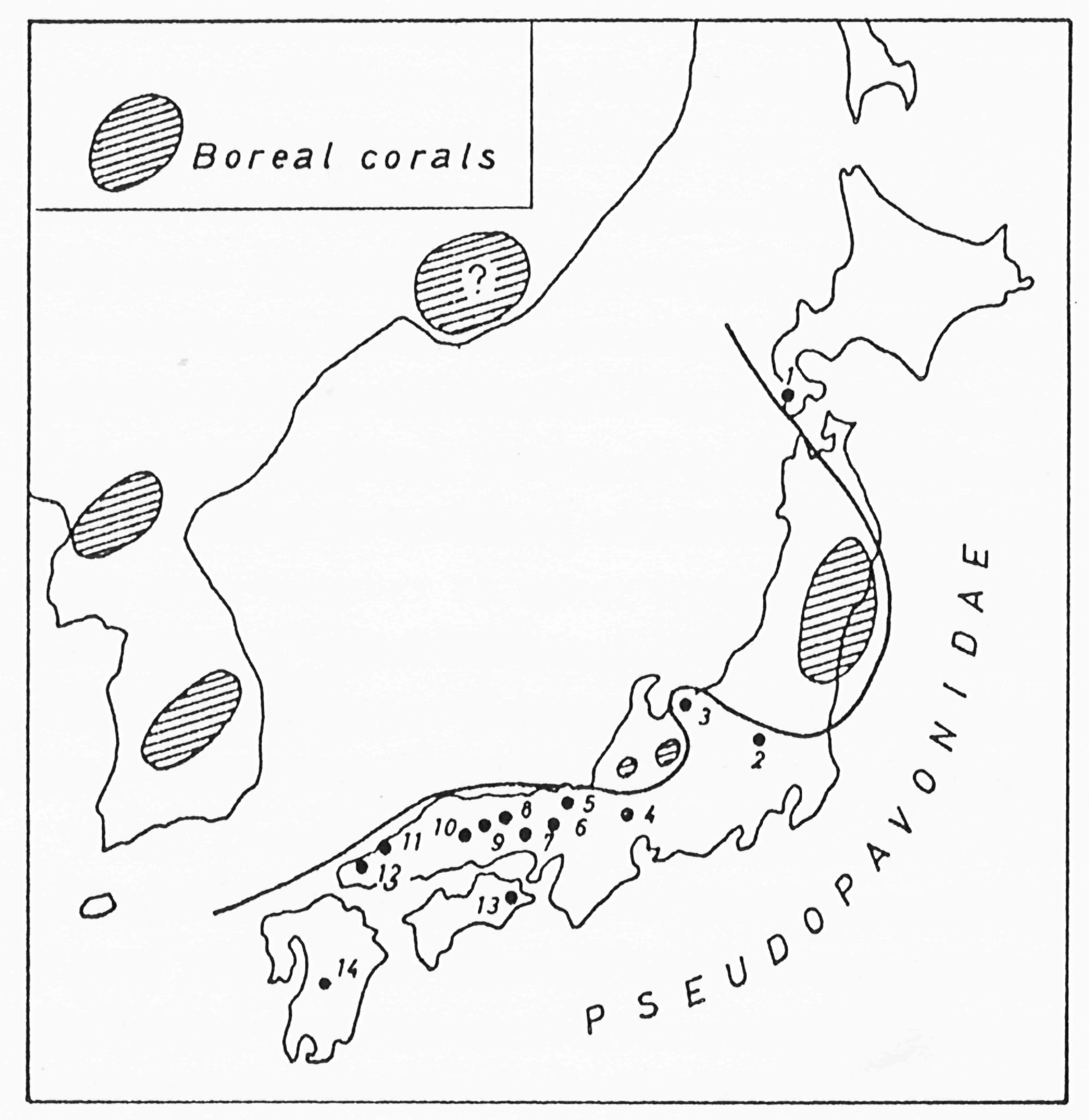
テチス域におけるペルムにおける四射サンゴの絶滅様式(Ezaki, 1997).生息環境の変化に応じた形態的・系統的な序列が明瞭である.
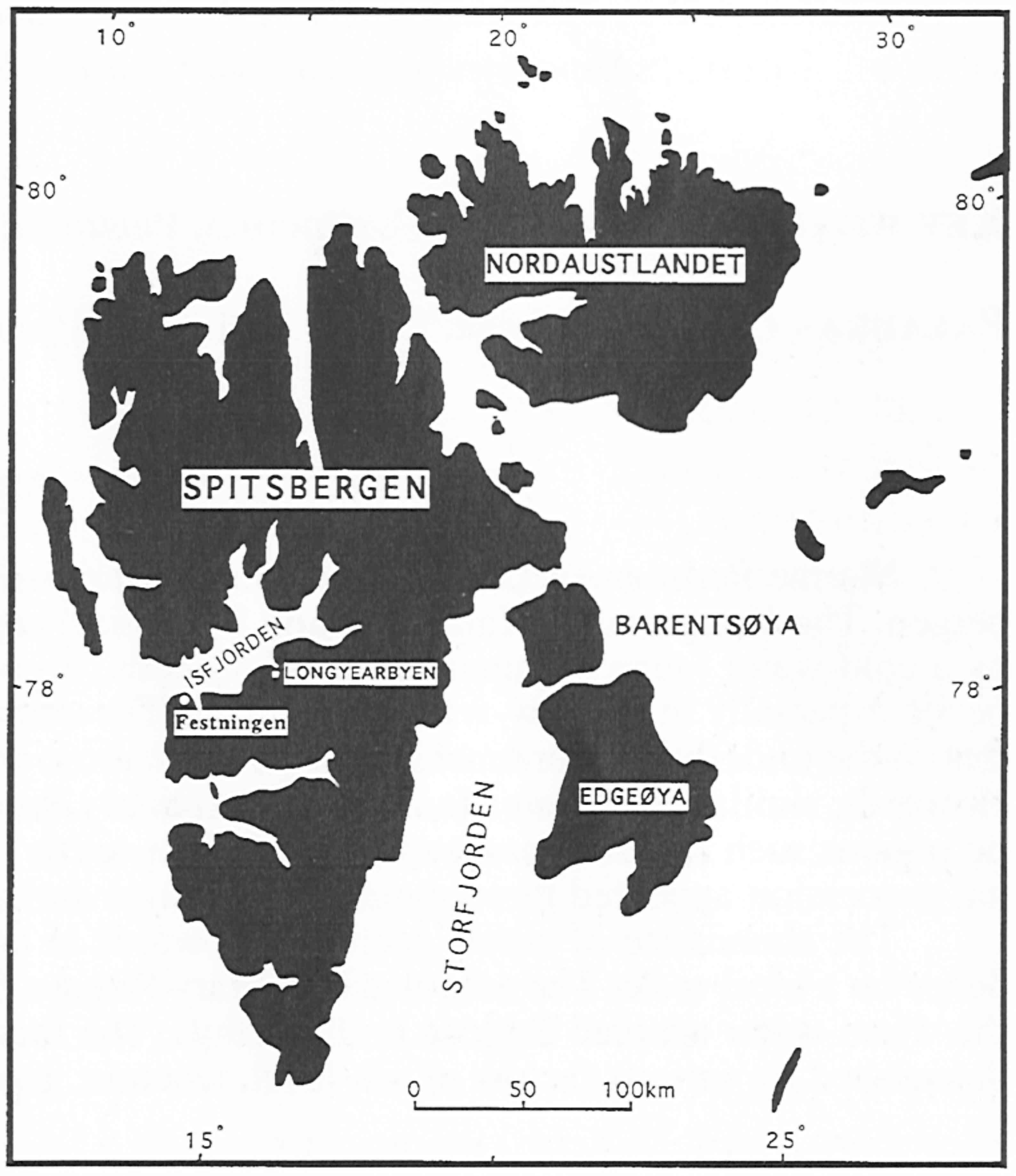

ペルム紀ボレアル域のサンゴ.(左)スピッツベルゲンはペルム紀にボレアル域に位置した.Festningenはペルム系Kapp Starostin層の模式地.(右)Kapp Starostin層から産出する代表的な四射サンゴ:1. Euryphyllum sp. A, 2, 3. Euryphyllum sp. B, 4, 5. Allotropiochisma svalbardicum, 6. Sassendalia turgidiseptata.(Ezaki, 1997).冷温性サンゴが生息していた.

(左)中生代最古級(アニシアン)のサンゴが産する(貴州省青岩).(右)中生代最古級(アニシアン)の群体六射サンゴ(中国貴州省青岩)


鳥巣式石灰岩中の六射サンゴ(上部ジュラ系:和歌山県湯浅)

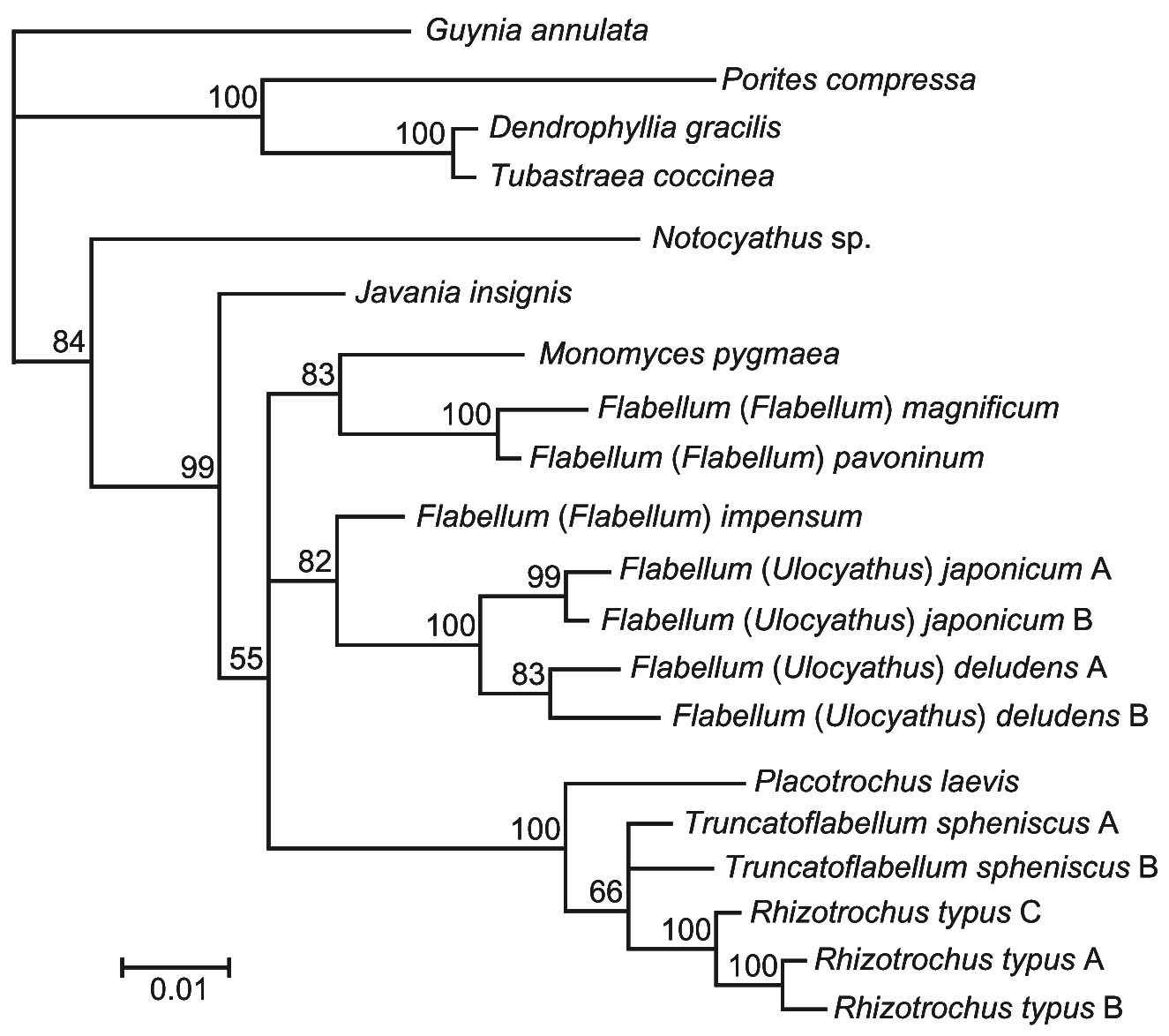
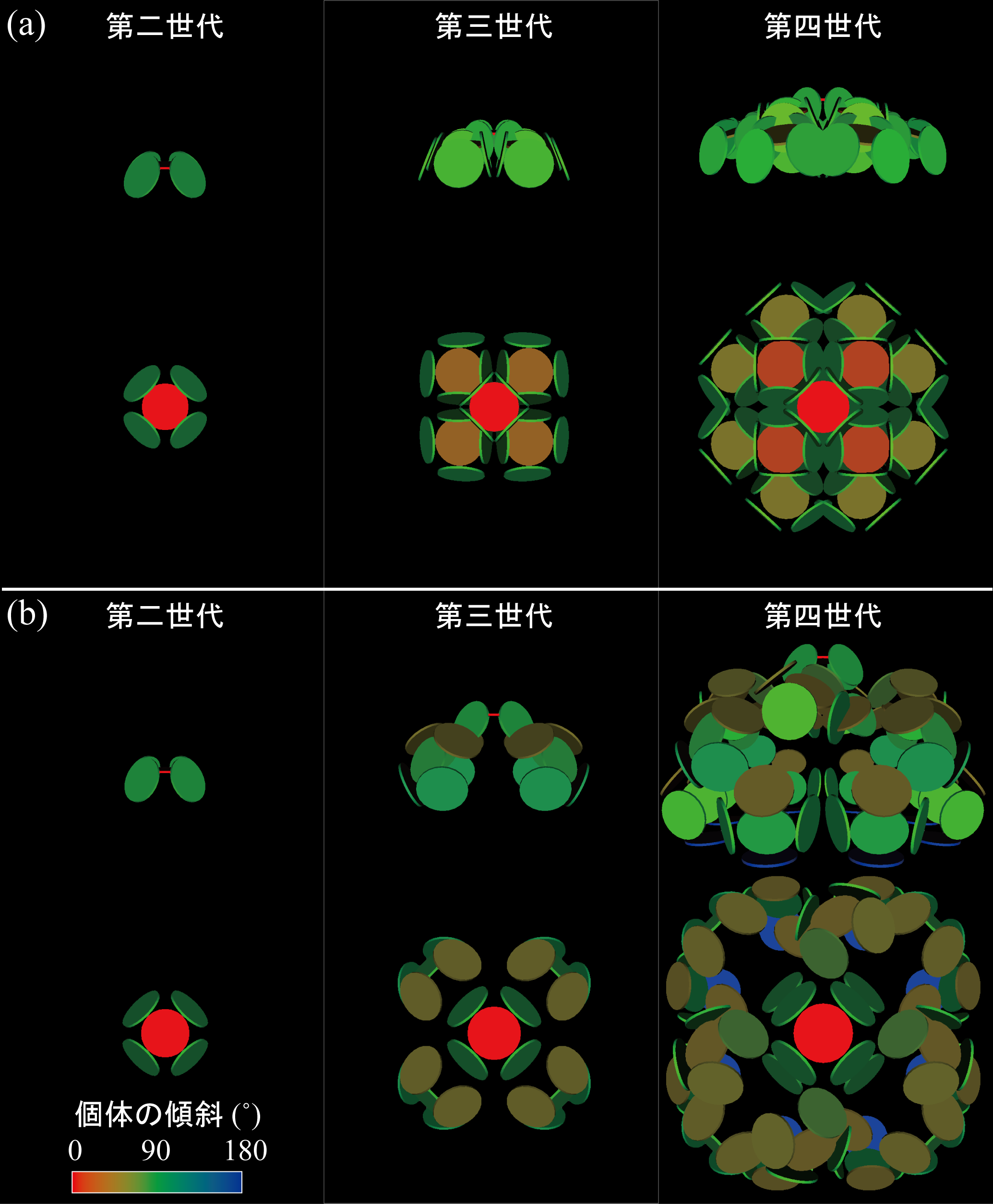
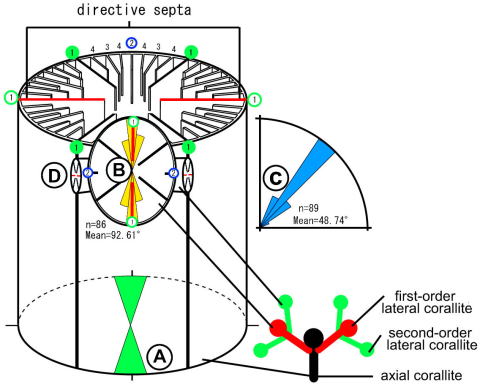
(左)各センスガイ科六射サンゴの外形(Tokuda et al., 2010).(右)16S rDNA と 28S rDNAデータに基づくセンスガイ科六射サンゴの系統関係(Tokuda et al., 2010)
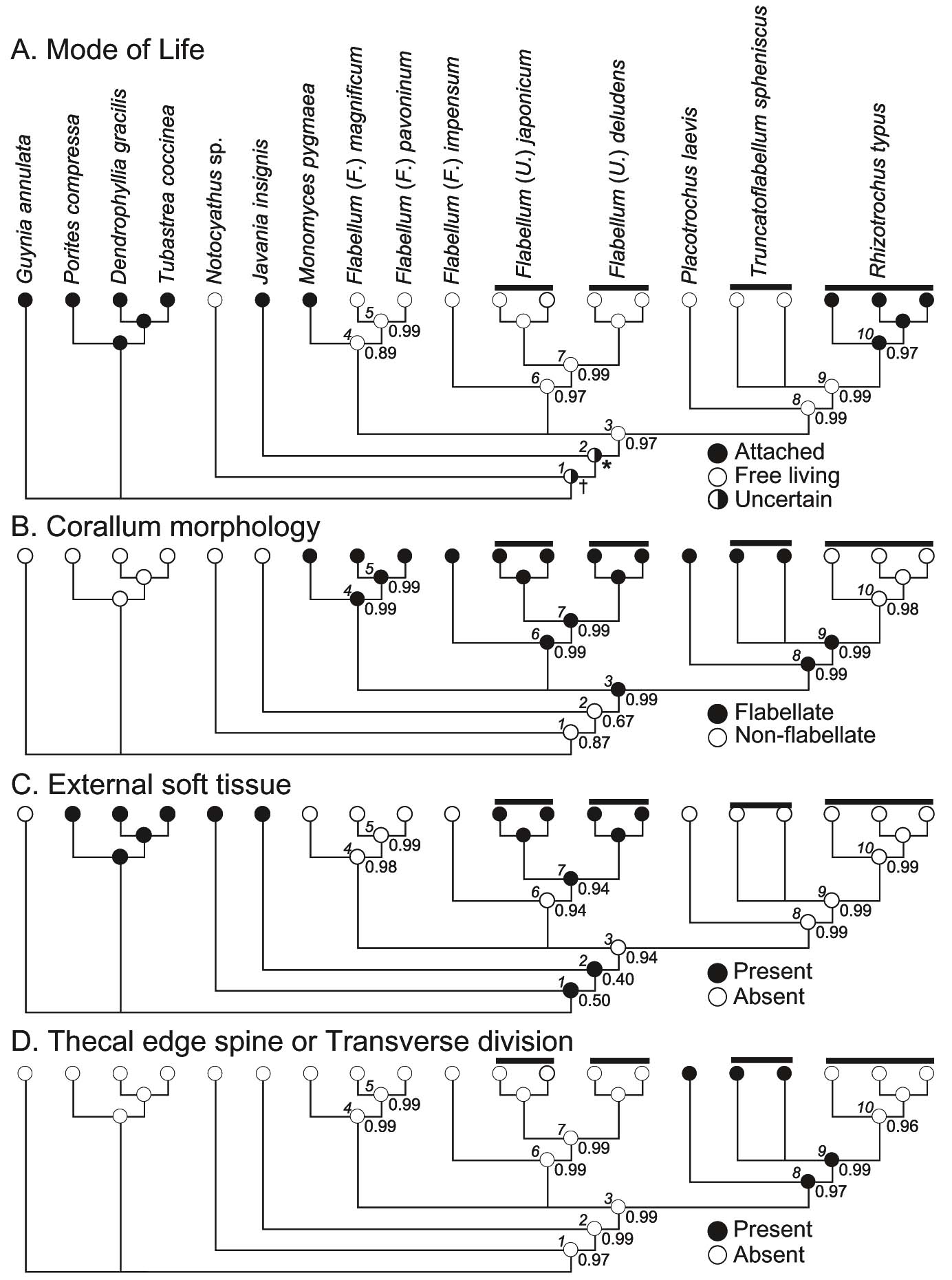
センスガイ科六射サンゴの各種形質に関する祖先形質解析(Tokuda et al., 2010)
無性増殖・成長様式の規則性と時代的変遷(多様化)・群体性生物の理論形態解析

キサンゴ科六射サンゴの多様な成長形態.「内因的な出芽の規則性」のもとで,どのように群体形態を変えているのか.生息環境の違いに呼応した巧妙な成長戦略が読み取れる.写真提供:千徳明日香博士
「如何にモジュールが増え,如何に個が存続,自らが環境を創出してきたのか」,「系統的・発生的な形態規制の意味内容」,「如何に成長型が形成,選択されてきたのか」,「群体性生物のかたちづくり」,「無性増殖の規則性の解明」,「群体サンゴ成長型のコンピューターシミュレーションと環境要因との関連」


(左)後期デボン紀四射サンゴの風化表面で出芽が頻繁に生じている(ロシア ウスチ-カタフ Ust-Katav層)(右)中期デボン紀四射サンゴの風化表面で4分裂が頻繁に生じている(ドイツ アイフェル Ermberg層)
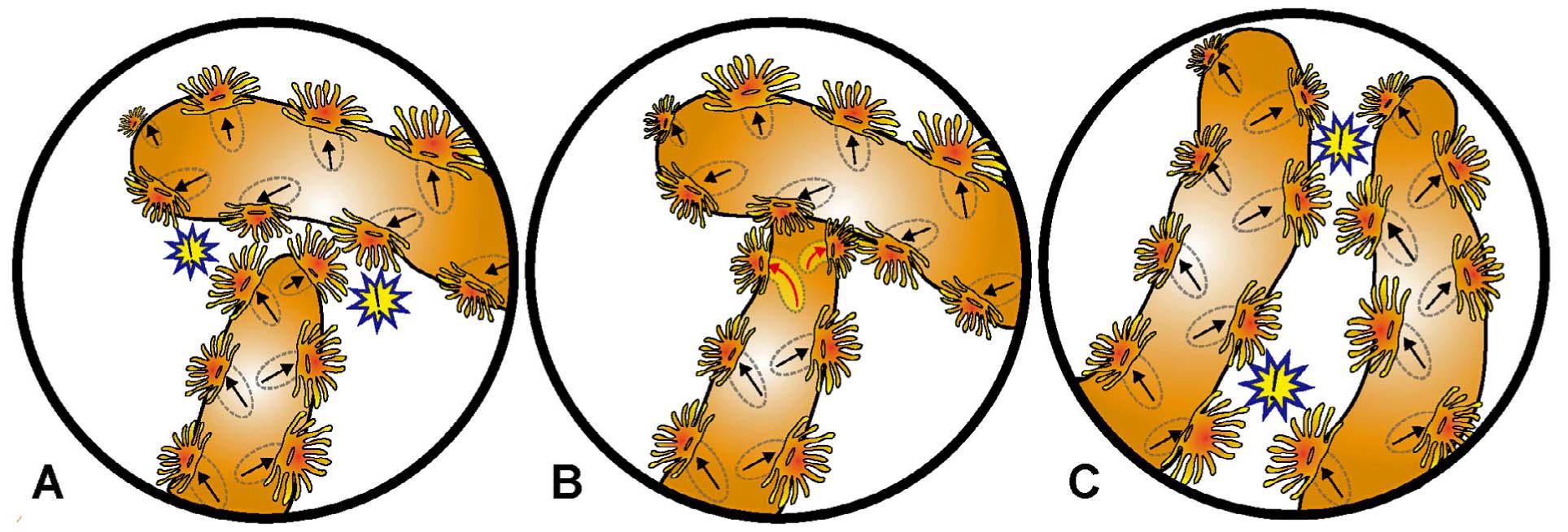
「無性増殖様式」と「群体形成様式」の相互関連性.サンゴの分裂様式には,規則性が内包されている.
.jpg)
Stauria favosaにみられる4分裂のプロセス(Ezaki & Yasuhata, 2004)
-14.jpg)
(左)Stauria favosaにみられる分裂の規則性(Ezaki & Yasuhata, 2004).分裂に際し,極性を維持した原隔壁が形成され,親個体と娘個体の極性は必ず堅持される.
.jpg)
Stauria favosaにみられる回春現象と4分裂のプロセス(Ezaki & Yasuhata, 2005)
.jpg)
Stauria favosaにみられる回春現象の影響を受けた4分裂の模式図(Ezaki & Yasuhata, 2005).分裂に際して,先ず特定の極性を維持した4枚の原隔壁が形成される.
Yamatophyllum ultimumに特徴的な2分裂(Ezaki & Kato, 2014).分裂後,親の個体と同じ方向に主隔壁(白丸)が形成される.
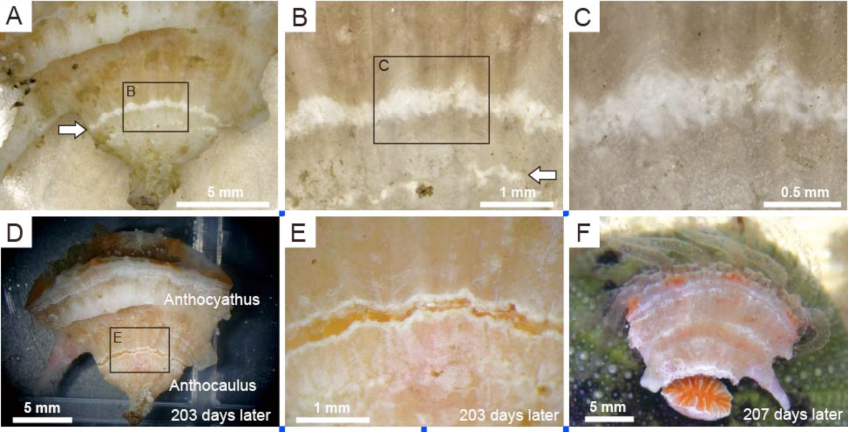
Truncatoflabellum spheniscusのanthocaulusに特徴的な横分裂のための脱石灰(白色帯部)(Tokuda et al., 2017)

Truncatoflabellum spheniscusの脱石灰後の軟体部の横分裂(Tokuda et al., 2017).分裂直後の軟体部が特徴的.
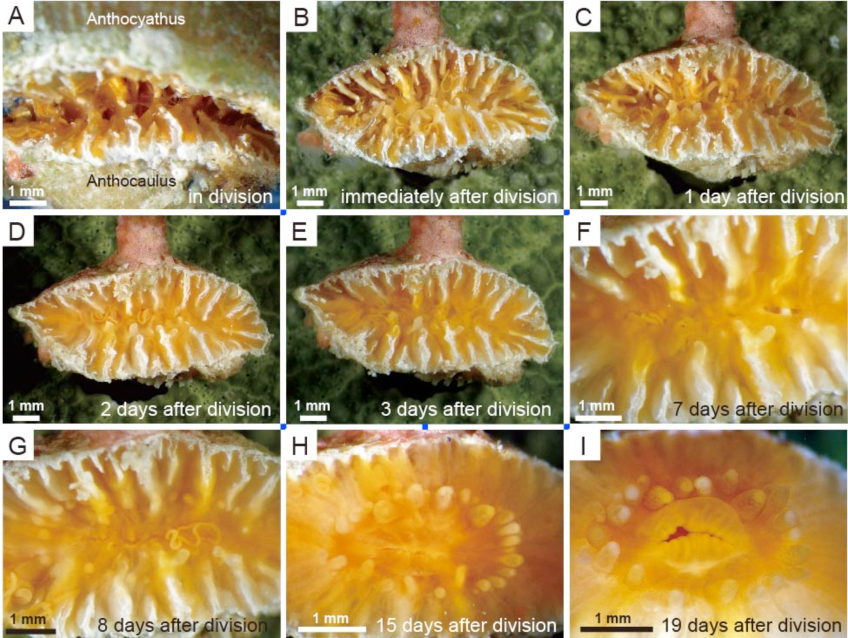
Truncatoflabellum spheniscusの横分裂後のanthocaulusのポリプの再生(Tokuda et al., 2017)早急に触手や口が再生される.
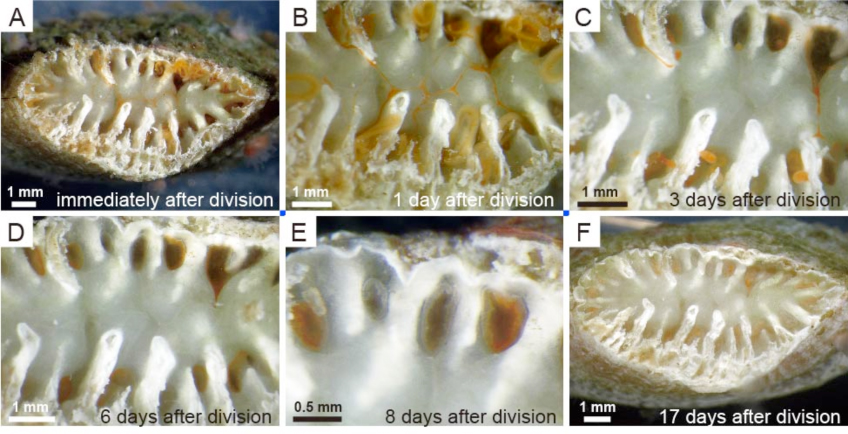
Truncatoflabellum spheniscusの横分裂後のanthocyathusの分裂部の修復(Tokuda et al., 2017)

Truncatoflabellum spheniscusの横分裂後の骨格の再成長と成長率(Tokuda et al., 2017).横分裂に先立ち側方にスパインが形成される.
「無性増殖様式」と「群体形成様式」の相互関連性.サンゴの出芽様式には,規則性が内包されている.
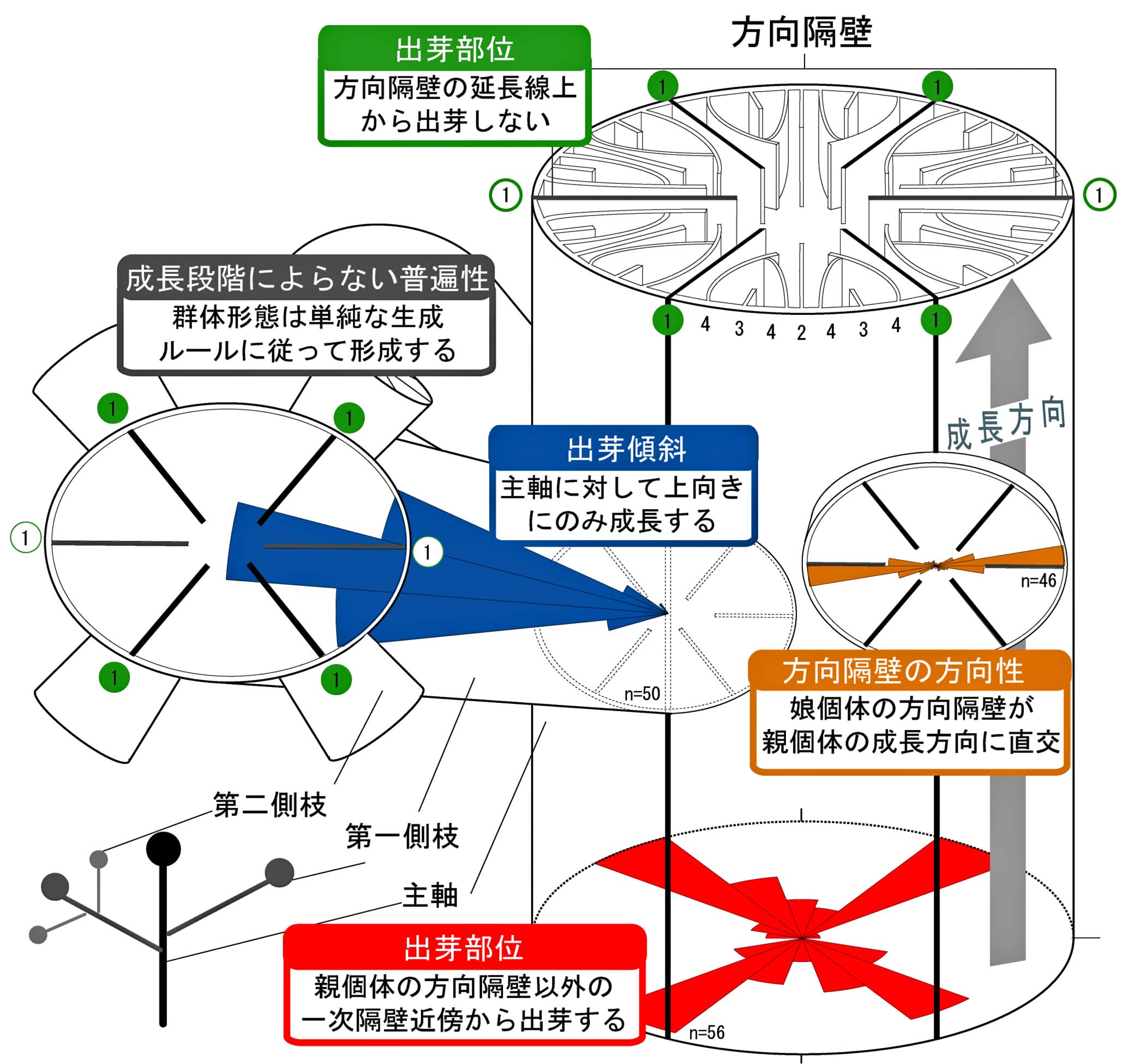
-1.jpg)
(上)非造礁性枝状六射サンゴDendrophyllia arbusculaにおける出芽時の規則性(Sentoku and Ezaki, 2012b).(下)塊状六射サンゴTubastraea coccineaと枝状サンゴDendrophyllia arbusculaとの比較概念図(Sentoku and Ezaki, 2012b).枝状と塊状に関わらず出芽は同じ規則性に従っている.
(a)Tubastraea coccineaと(b)Dendrophyllia arbusculaにおける出芽時の規則性(Sentoku and Ezaki, 2012b). 規則性は側枝の世代を超えて成り立っている.

Tubastraea coccineaの出芽様式(Sentoku and Ezaki, 2012b).塊状サンゴでも枝状サンゴDendrophyllia arbusculaと同様の規則性を示す.
-5.jpg)
-9.jpg)
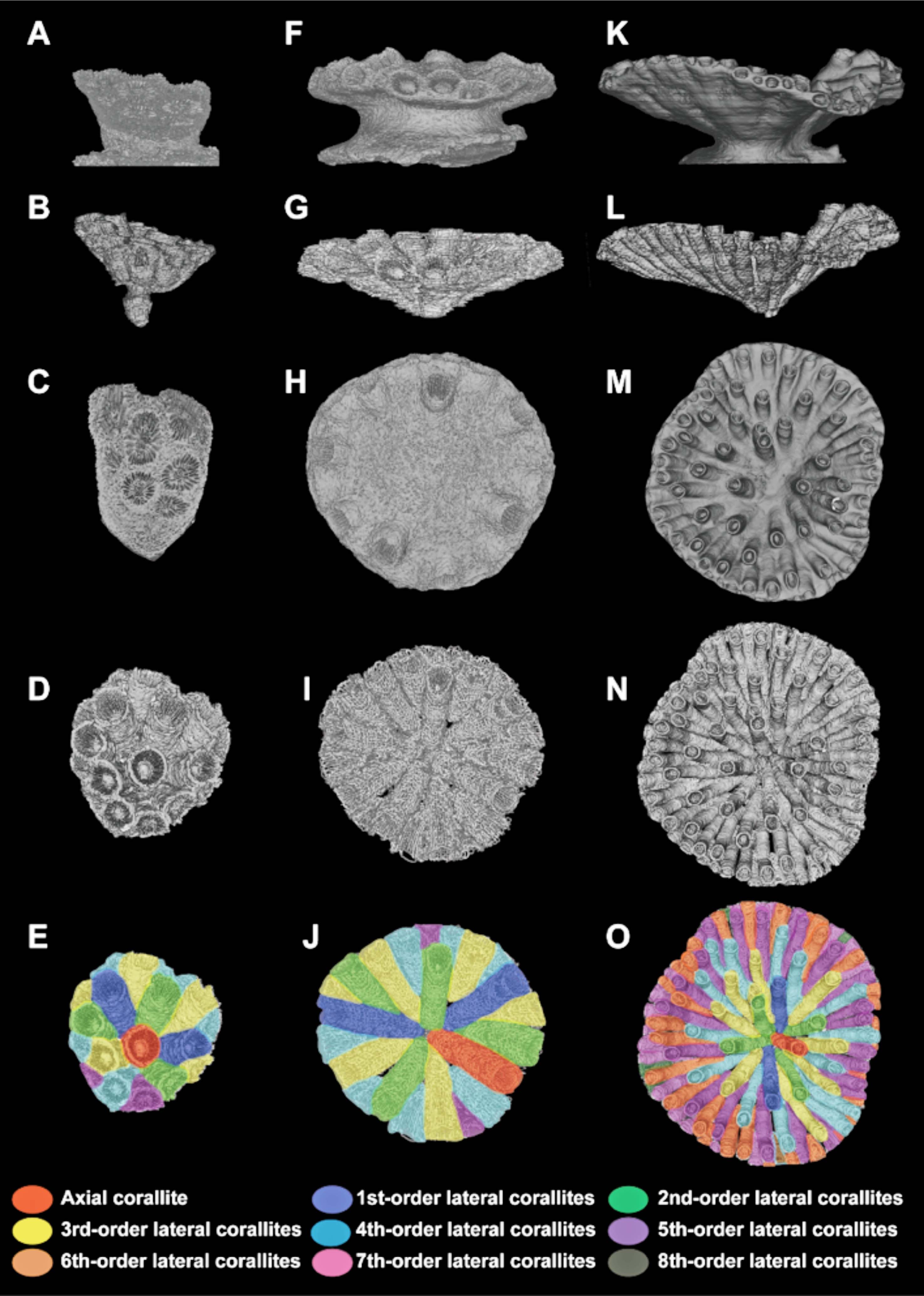
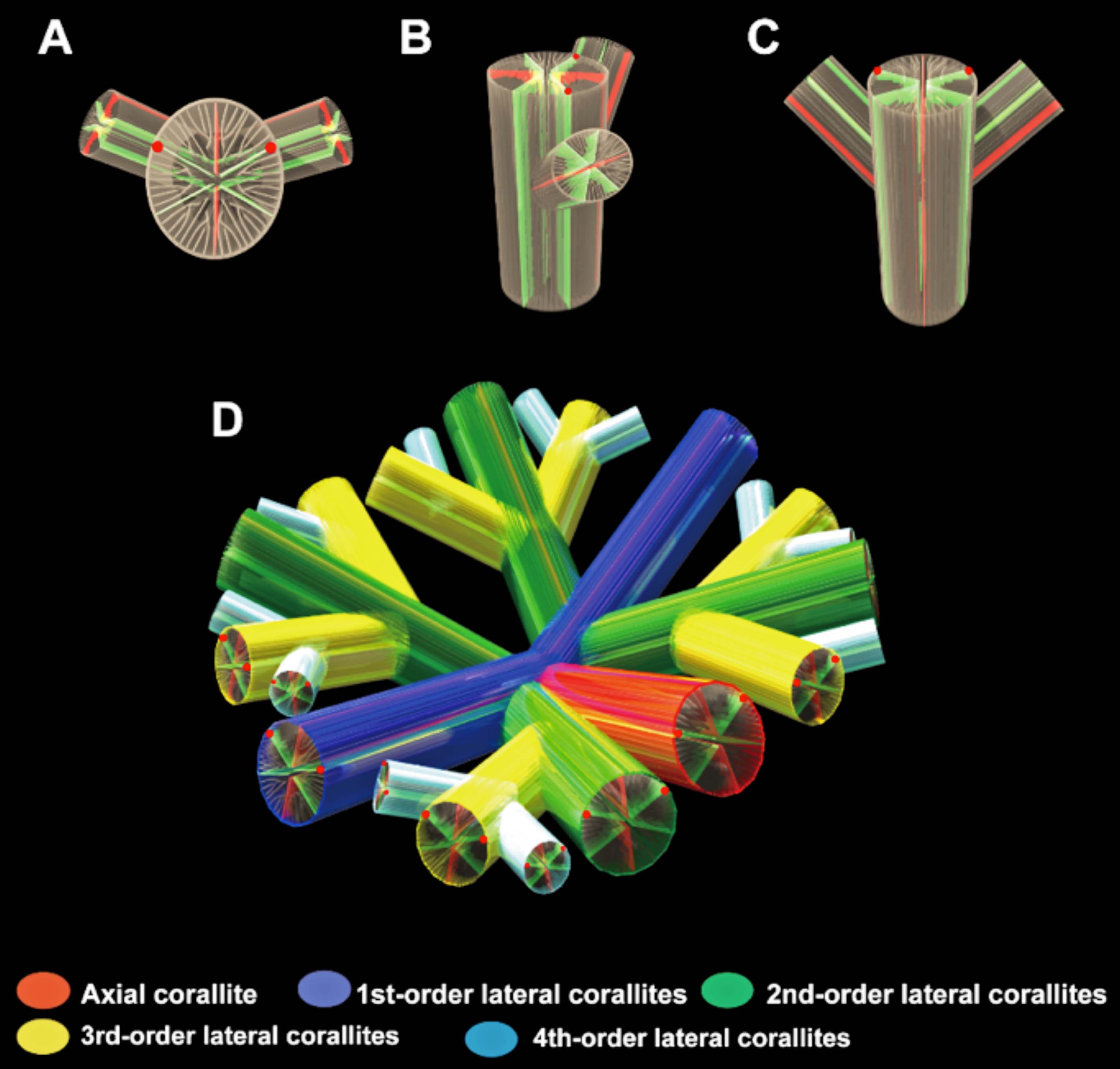
(左)理論モデルを構築する際に必要な3つのパラメーター(出芽傾斜,回転角,出芽間隔).(右)Tubastraea coccineaとDendrophyllia arbusculaのコンピューターシミュレーション図(A, C)と実際の群体骨格(B, D). スケール 1cm. (Ohno et al., 2015)


(左)出芽方位とその制約条件.娘個体の起点と親個体のX,Yによって定義される平面での断面.(a) 出芽方位と娘個体を形成する方向. (b), (c) 娘個体の初期径による,出芽方位φの制約. (b)出芽方位の最小値φmin.(c)出芽方位の最大値φmax.(右)処理の流れ.ステータスs(n)は個体nの成長の状態を示す (1: 通常成長,2: 初期成長,3: 他個体との干渉による成長停止,4: 底面との干渉による成長停止)(大野他, 2016)
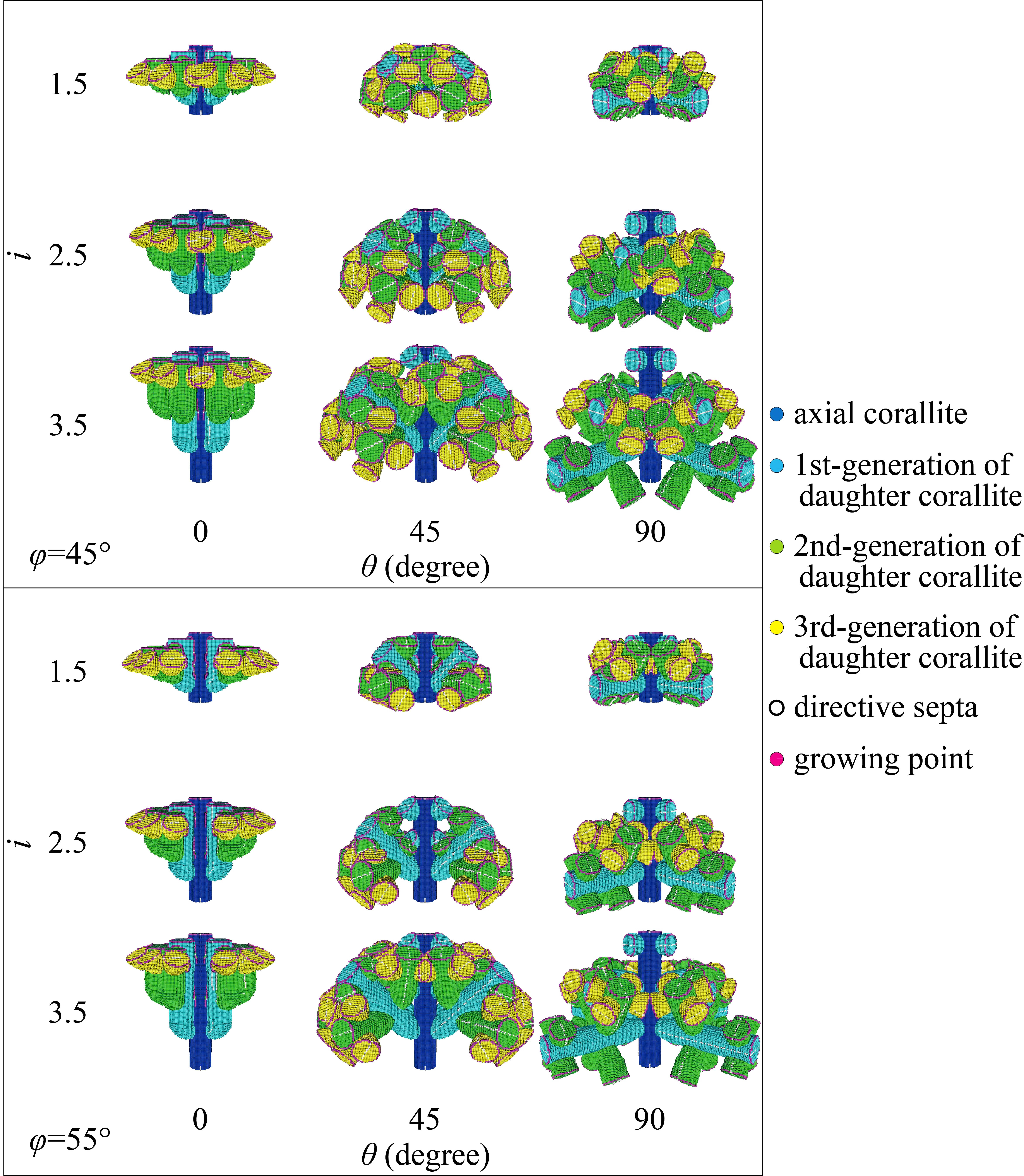
サンゴ群体の形態形成モデルで,3つのパラメーター(出芽傾斜,回転角,出芽間隔)を変化させたときの理論形態(Ohno et al., 2015)
群体内の各個体の成長方向が主軸の成長方向に対してなす角の可視化例. (a) θ = 10°,φ = 45°,i = 1.5,(b) θ = 65°,φ = 45°,i = 2.5(大野他, 2016)
-7.jpg)
特定のパラメーター値(出芽傾斜,回転角,出芽間隔)下での群体の体積と枝総数の時間的な変化(Ohno et al., 2015)
-8.jpg)
特定のパラメーター値(出芽傾斜,回転角,出芽間隔)下での枝個体数の時間的な変化(Ohno et al., 2015)
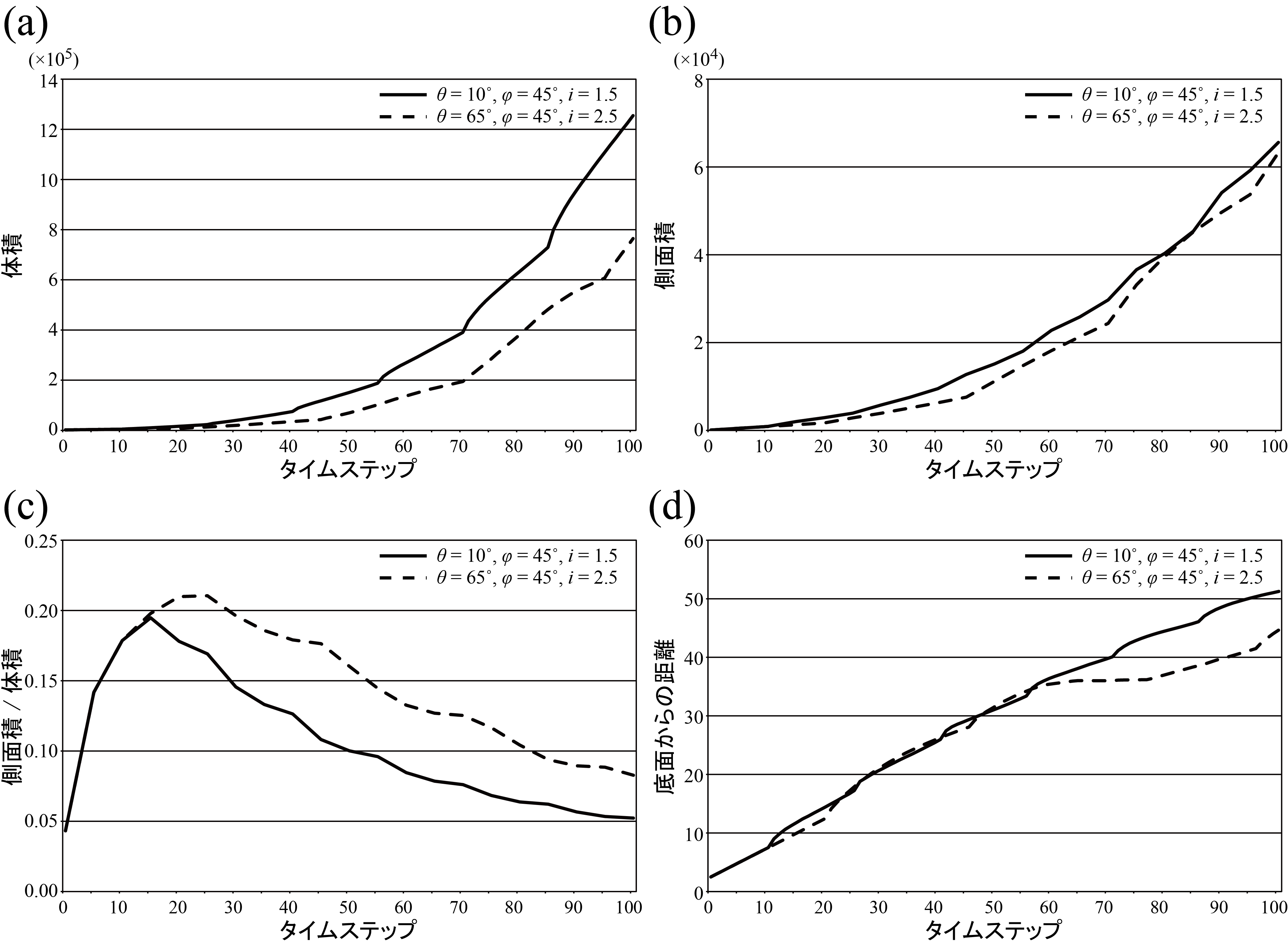
各パラメータをθ = 10°,φ = 45°,i = 1.5及びθ = 65°,φ = 45°,i = 2.5に設定して得られた群体の性質の時間変化.(a)群体の体積.(b)群体の投影面積.(c)群体の投影面積-体積比.(d)群体の重心位置(大野他, 2016)
(左)Dendrophyllia ehrenbergianaの出芽様式(Sentoku and Ezaki, 2012c).出芽の方向に極性が認められる.(右)Cyathelia axillarisの出芽様式(Sentoku and Ezaki, 2012d).方向隔壁近傍で出芽は生じないが,出芽個体の方向隔壁は親個体の成長方向に配置する.結果的に,効率的な成長・採餌空間を造り出している.

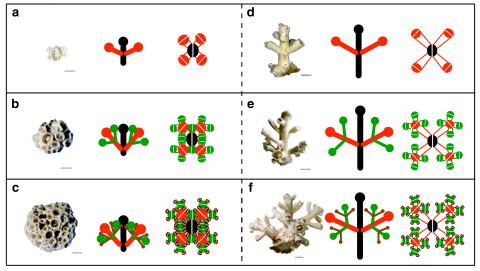
キサンゴ科で認められる出芽の規則性と適合範囲.A Dendrophyllia arbuscula, B D. ijimai, C Tubastraea micrantha , D T. coccinea, E D. ehrenbergiana, F Turbinaria peltata, G D. boschmai, H D. cribrosa, I Cyathelia axillatis. Scale = 1 cm.(千徳・江﨑, 2016)
-2.jpg)
Dendrophyllia boschmai (A-D)とD. cribrosa (E-H)の形態(Sentoku and Ezaki, 2013)
-4.jpg)
Dendrophyllia boschmai (A-E)とD. cribrosa (H-M)で認められる出芽様式(Sentoku and Ezaki, 2013)
-8.jpg)
Dendrophyllia boschmai (A-D)とD. cribrosa (E-H)で認められる出芽様式と群体形成様式(Sentoku and Ezaki, 2013).規則的なかつ巧みな方法でそれぞれの成長形態を造り出している.

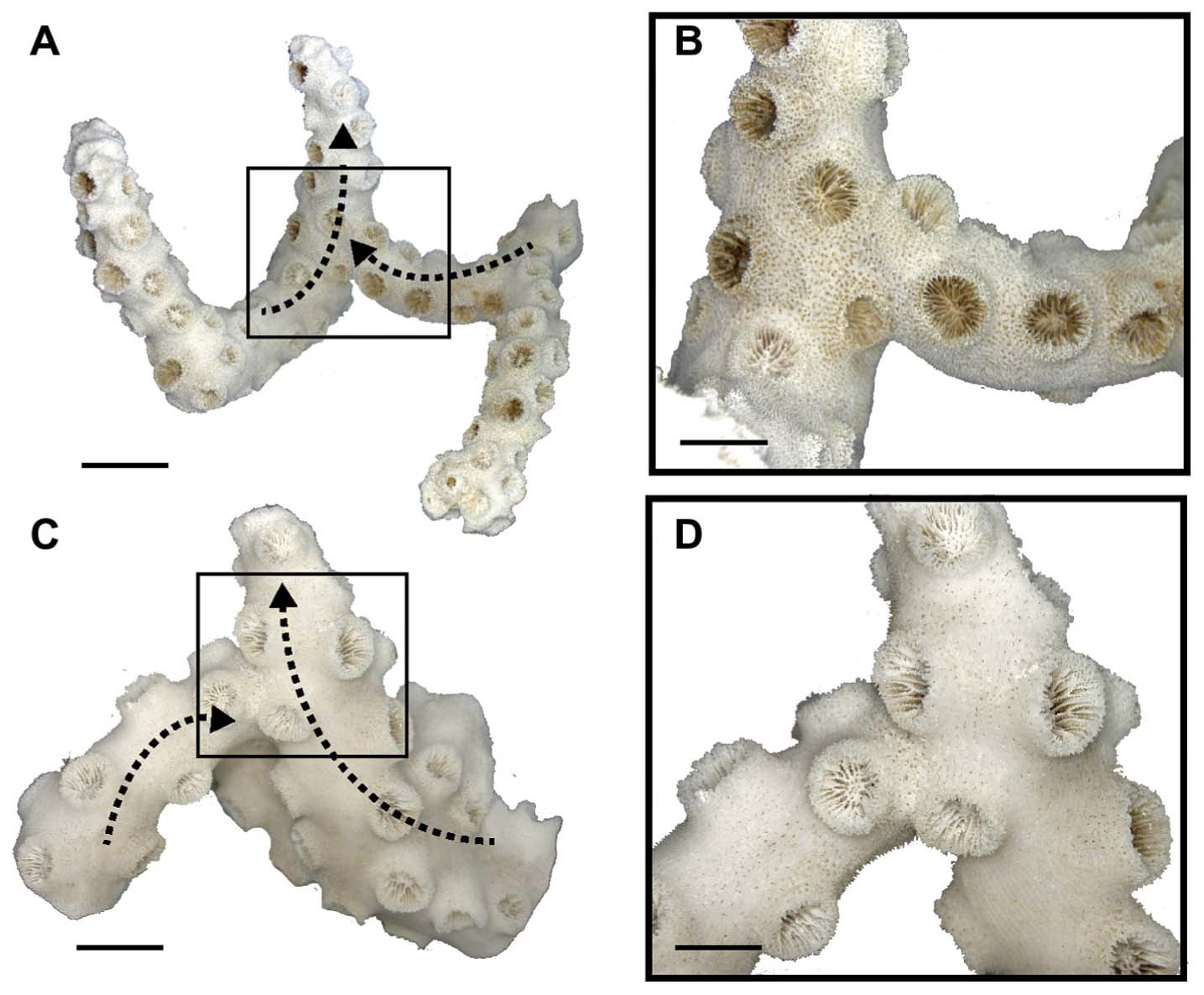
Dendrophyllia cribrosaで認められる枝の癒合部分(Sentoku et al., 2015)
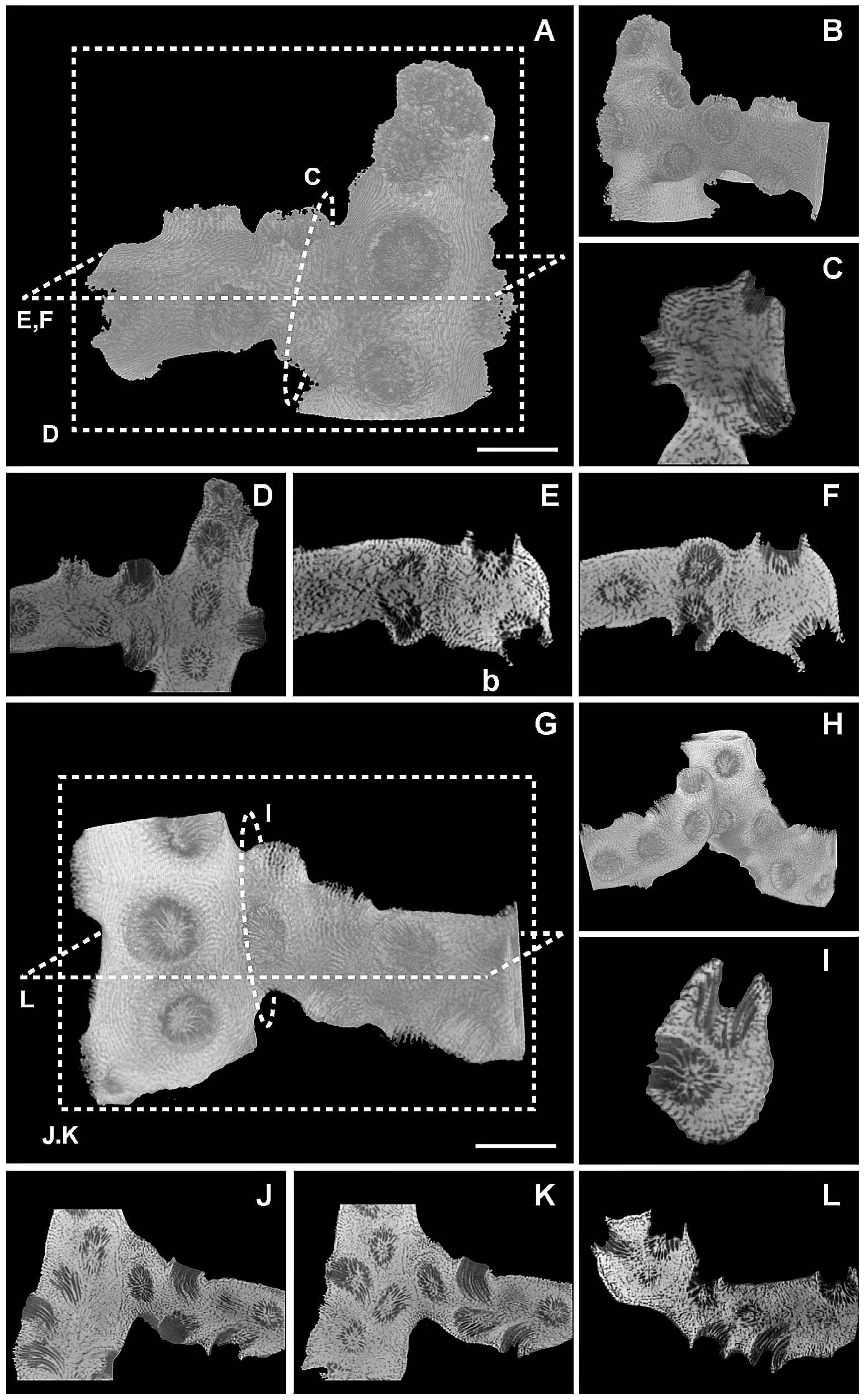
Dendrophyllia cribrosaで認められる枝の癒合部分のX線CT画像(Sentoku et al., 2015)
Dendrophyllia cribrosaで認められる枝の癒合部分..近接個体間は触手を通じて互いに他を認識し,衝突を事前に回避している.(Sentoku et al., 2015)
Turbinaria peltataで見られる世代ごとの出芽個体の分布. X線CT画像から解析(Sentoku et al., 2015)
Turbinaria peltataで見られる世代ごとの出芽様式の模式図(Sentoku et al., 2015).共骨を取り除くと,各個体は出芽規則に従い形成されていることがわかる.

Cyathelia axillarisの骨格の概要(千徳, 2013)
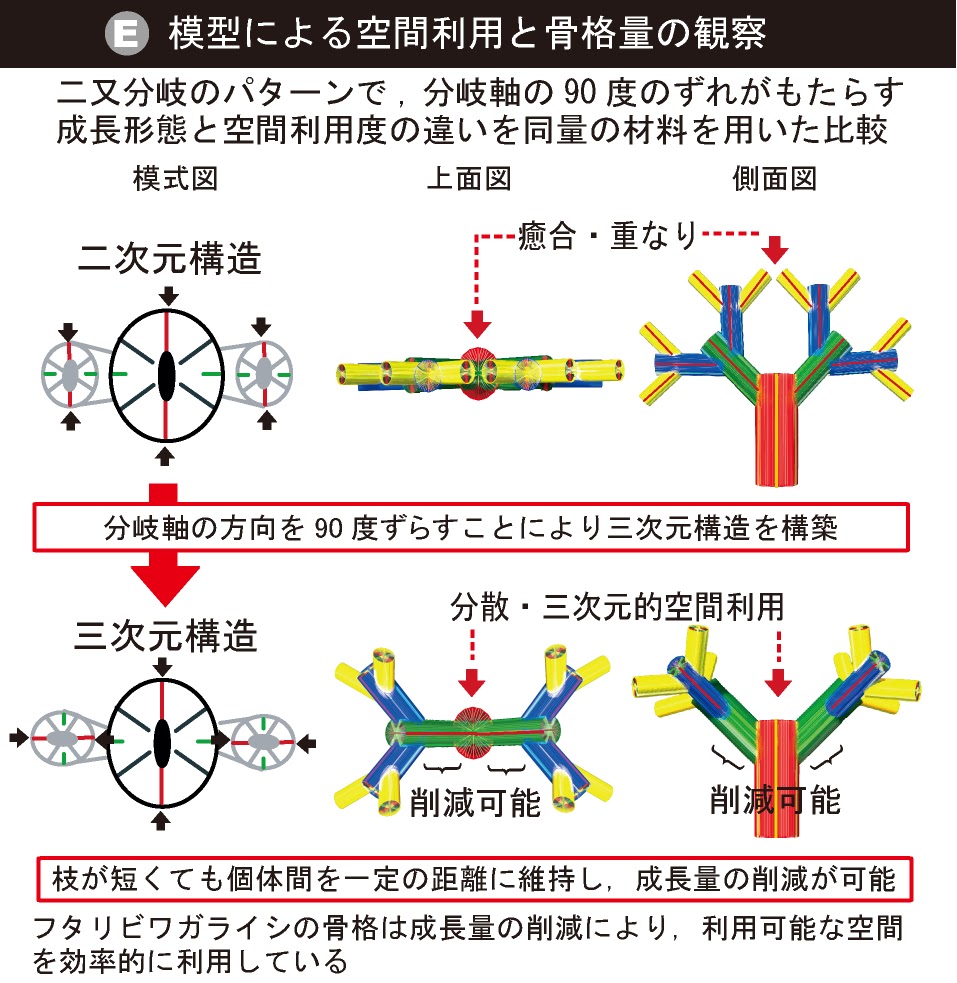
Cyathelia axillarisにおける成長空間の効率的な活用(千徳, 2013)
個体・群体の存在(集合・統合)様式と生活史戦略
「個体とは」,「群体とは」,「個体と群体の構造的・機能的な関係」,「個は如何に造られ,創られてきたのか」,「各骨格構成要素の在り様(生態的・構造的な存在理由)」

四射サンゴにおける隔壁の分類と対応関係.「Catasepta」(Ezaki, 1989)の概念を導入することにより,隔壁の記載的な分類と成因的な分類の関係や,隔壁の挿入様式のより正確な解析が可能になる.

サンゴ形態の3大基本要素(壁,隔壁,床板)とそれらの派生要素


単体四射サンゴDokophyllumの個体間をつなぐ連結細管(connecting tubule).スケール:1cm(シルル系:スウェーデン ゴトランド島).



四射サンゴMicrocyclus sp.で認められる明瞭な左右相称性と極性.主隔壁溝(cardinal fossula)の発達が顕著(中部デボン系[Eifel階]Ahrdorf層:ドイツ).


四射サンゴAnkhelasma typicumで認められる明瞭な左右相称性と極性.主隔壁溝(cardinal fossula)の発達が顕著(上部石炭系:北米ユタ州).



六射サンゴで認められる隔壁の“放射相称的”配列.(左)Microphyllia sp.(上部ジュラ系:ドイツ Nattheim, Baden Württemberg).(中)Caryophyllidサンゴ(和歌山県南部).(右)Caryophyllidサンゴ(和歌山県南部).
.jpg)
.jpg)
.jpg)
六射サンゴAlatotrochusで認められる“扁平の”個体形態(沖永良部島沖).(左)個体側面(肋の発達が顕著.個体下部がくさび状).(中)莢部内の隔壁配列.(右)隔壁の張り出し.


サンゴ化石が選択的にドロマイト化作用を受けたことで,形態の基本要素が顕著に認められる(下部ペルム系:ロシア ステルリタマク)


サンゴ化石が選択的にドロマイト化作用を受けたことで,形態の基本要素が顕著に認められる(下部ペルム系:ロシア ステルリタマク)

サンゴ化石が選択的に珪化作用を受けたことで,骨格表面の特徴を把握しやすい.スケール 1mm(上部ジュラ系:ドイツ Nattheim, Baden Württemberg)


化石の保存状態が良く骨格表面の特徴を把握しやすい.スケール 1mm(上部白亜系:Gosau, Austria)

床板・四射サンゴの成長様式の時代変遷(Scrutton, 1988を改変;江﨑, 2004)
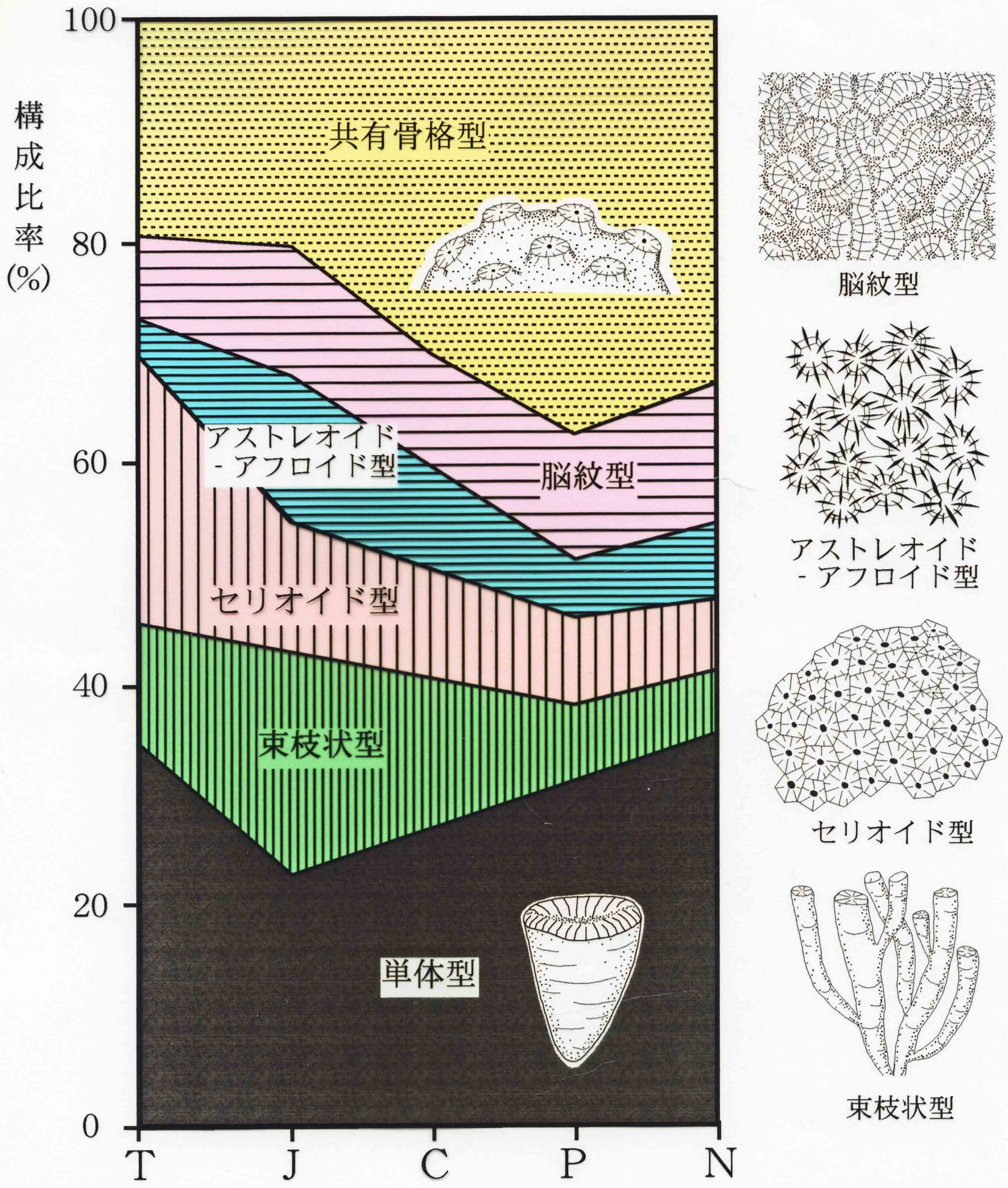
六射サンゴの成長形態の時代変遷(Wood, 1999を改変;江﨑, 2004)


Fungiaで見られる無性増殖.(左)口盤側の軟体部.(右)柄部(anthocaulus)から出芽個体(anthocyathus)が成長途上である.


(左)瀕死のCycloserisの親個体表面から一斉に出芽した娘個体.スケール1cm.(右)瀕死のFungiaの親個体裏面から一斉に出芽した娘個体


Fungiaで見られる無性増殖.(左)まだ固着している2個体が隣り合っている.左側の個体の周縁部が,隣接サンゴ個体の存在により歪んでいる.(右)柄部(anthocaulus)から出芽個体(anthocyathus)が横分裂しかけている.


Fungiaで見られる無性増殖.(左)自由生活性サンゴがまだ固着している.個体の周縁部が,隣接藻類骨格の存在により歪んでいる.(右)柄部(anthocaulus)から出芽個体(anthocyathus)が成長している.


単体六射サンゴCaryophylliaの骨格表面に残された修復跡.
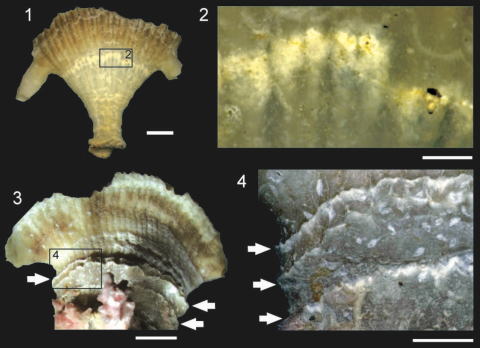
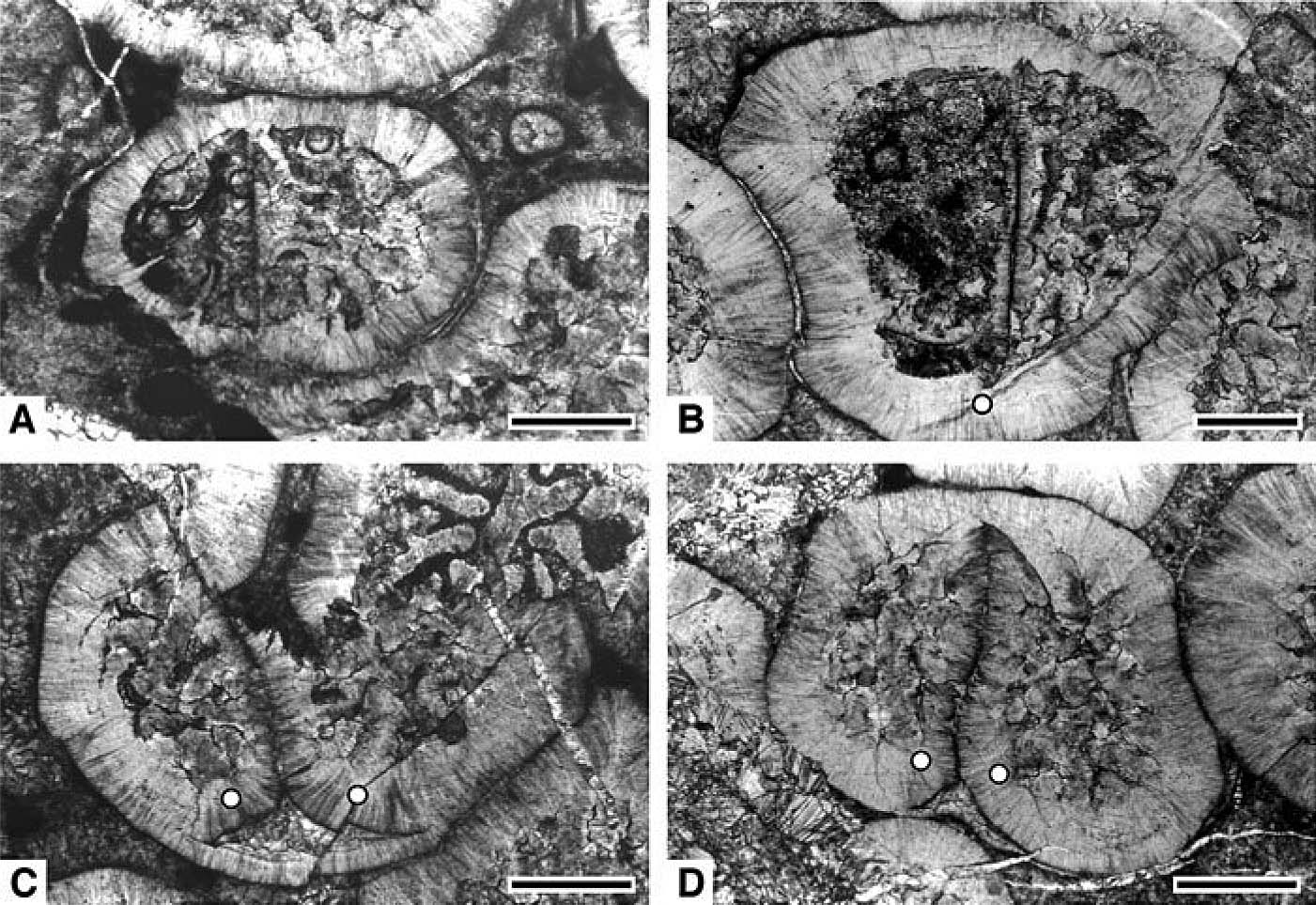
化石Truncatoflabellumに認められる無性増殖(横分裂)の痕跡(1,2)と分裂に起因する回春現象(3,4)(Tokuda and Ezaki, 2012)
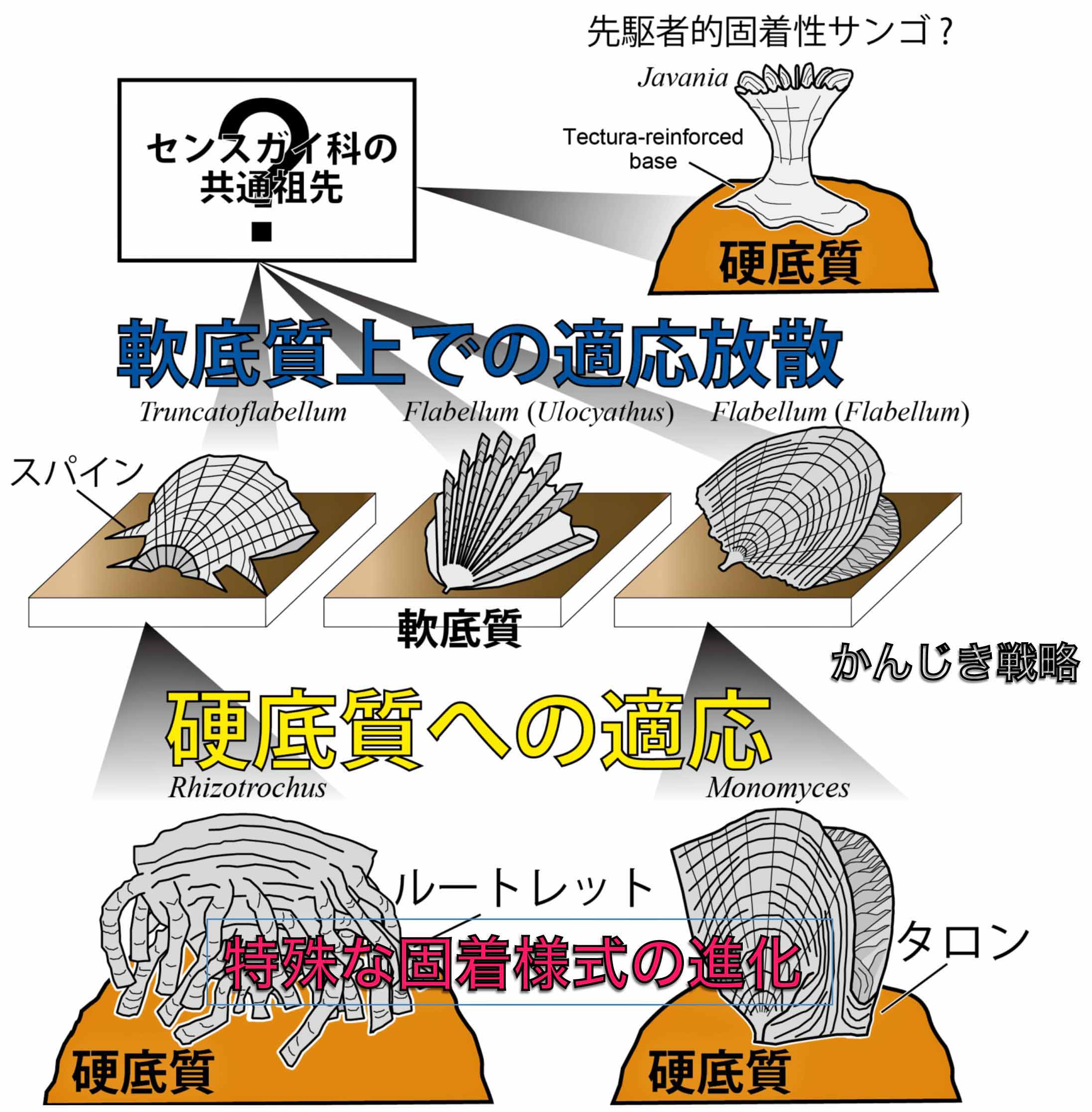
センスガイ科六射サンゴにみられる底質への適応戦略史(Tokuda and Ezaki, 2013).底質条件の変化に応じて生活様式を巧みに変化させている.
-7.jpg)
カンブリア紀サンゴCambroctoconus orientalisの隠棲環境での逆さ成長を示す風化面や研磨面(Ezaki et al., in press).開口部が下方を向いている場合が多い.中国山東省(張夏層)
-8.jpg)
カンブリア紀サンゴCambroctoconus orientalisの隠棲環境での生活様式,床板,固着器官(Ezaki et al., in press).個体の基底部では堅牢な固着器官が発達している.中国山東省(張夏層)
-12.jpg)
カンブリア紀サンゴCambroctoconus orientalisの隠棲環境での生活様式の模式図(Ezaki et al., in press).利用可能な隠棲空間の拡がりの程度によって,個体の大きさや出芽の頻度が即時的に変化している.中国山東省(張夏層)

深海単体六射サンゴDeltocyathus orientalisの軟体部の上面と側面.骨格部の上面,底部と側面(Sentoku et al., 2016)
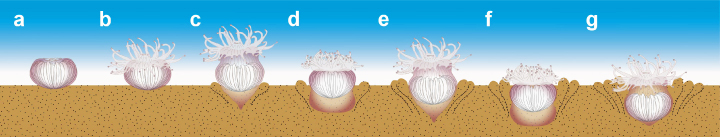
深海単体六射サンゴDeltocyathus orientalisの軟低質への潜行過程(Sentoku et al., 2016)
深海単体六射サンゴDeltocyathus orientalisでの能動的な上下移動のメカニズム.コステ内で海水を上下移動させ,軟体部を上部と下部で繰り返し膨縮させている(Sentoku et al., 2016)
骨格の成長記録保存能の理論的背景
「刺胞動物の生体リズムと骨格の分泌様式(『サンゴカレンダーの仕組み』)」,「サンゴ骨格の同位体・微量元素と環境指標性」,「サンゴ骨格の(超微細構造(『体のつくり』),「時計遺伝子の発現と生物鉱化作用」
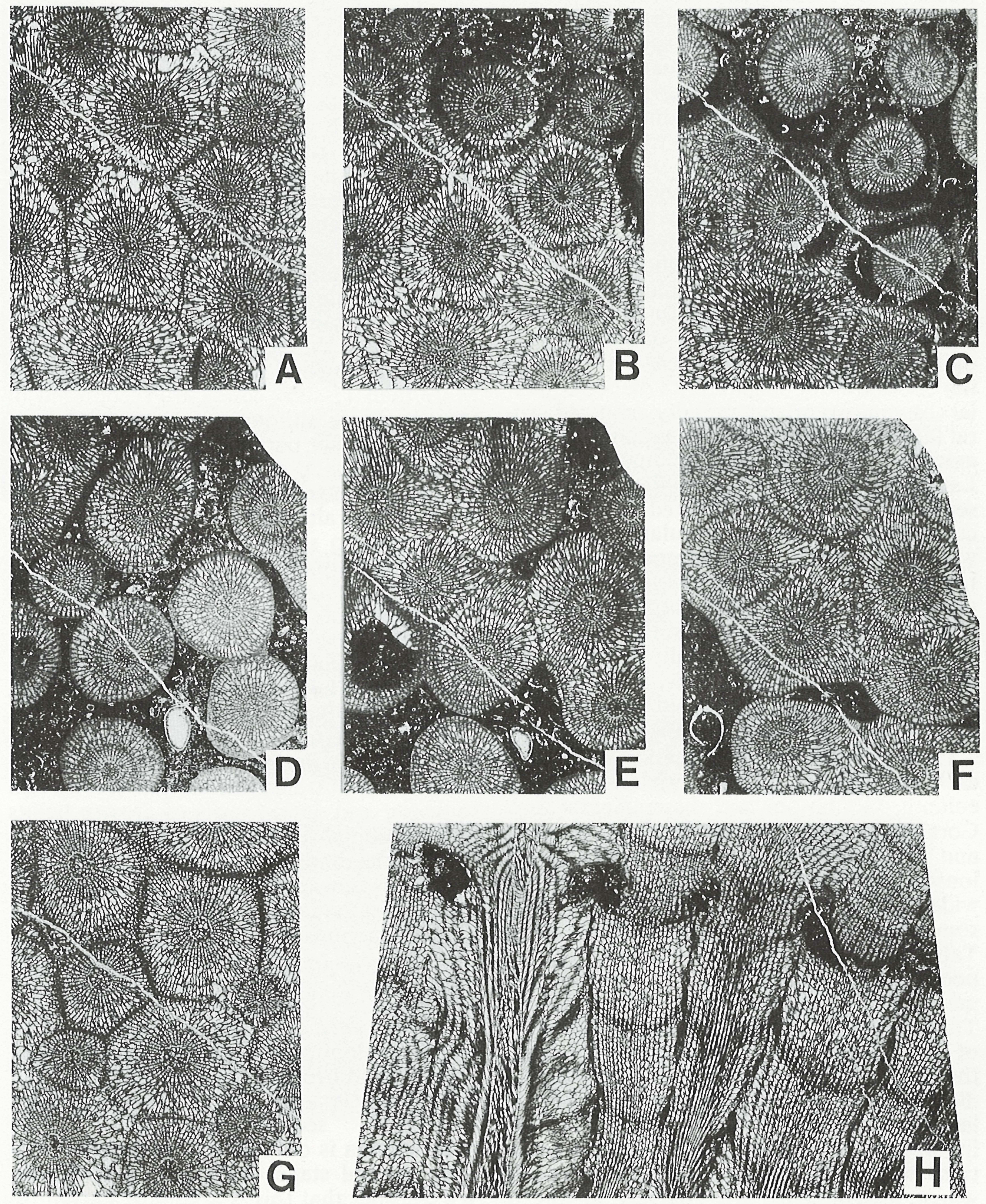
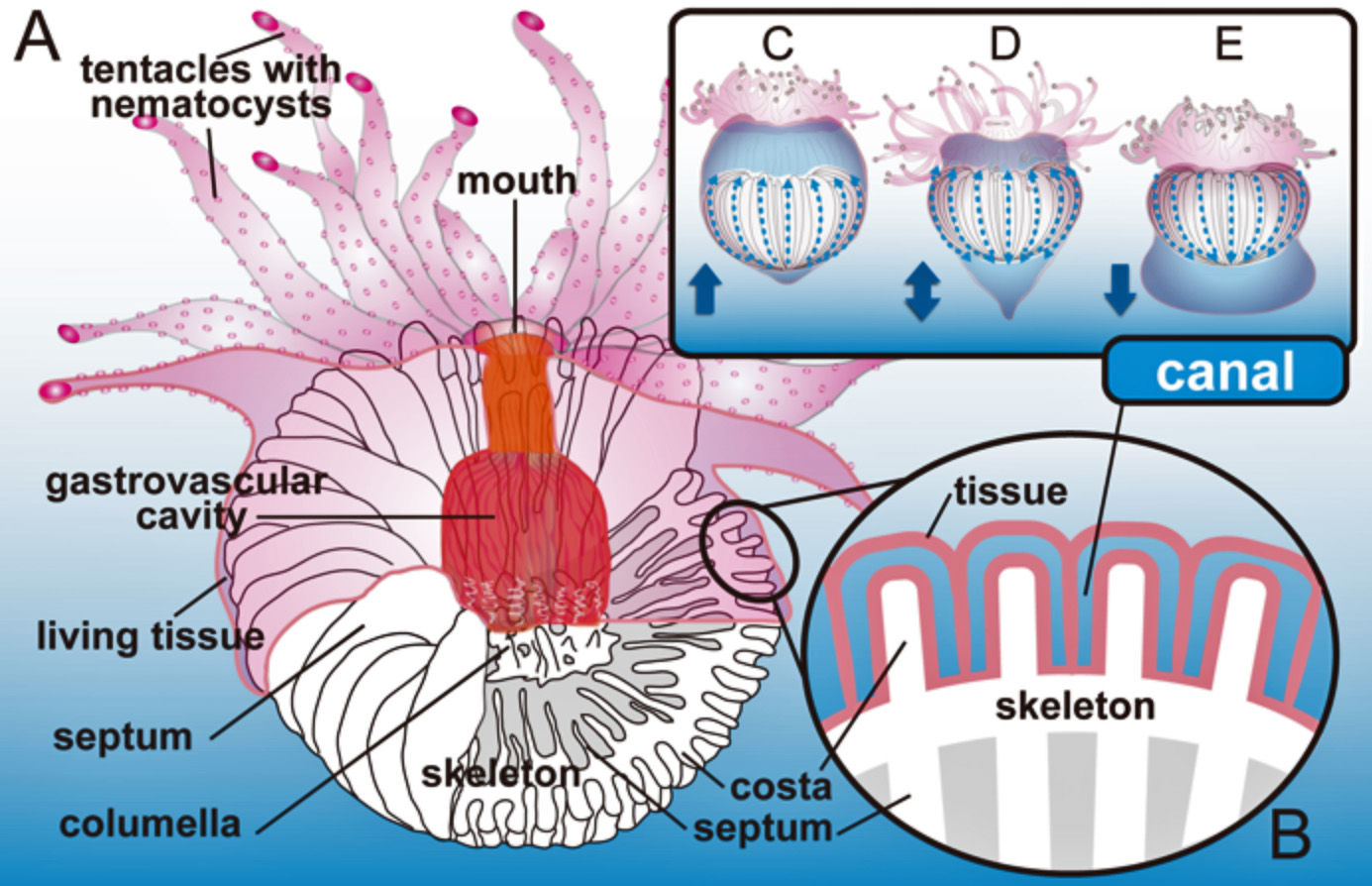
Wentzelella (W.) irregularisの連続横断面(Ezaki and Kato, 1989).回春により一時的に枝状の形状が形成される.

(左)Wentzelella (W.) irregularisの縦断面(Ezaki & Kato, 1989).年縞とストレスバンドが顕著.(中)Flabellum (F.) magnificumの骨格表面.様々な時間レベルの成長線が顕著.(右)F. (Ulocyathus) deludensの生体.周期的な生体反応を示す.

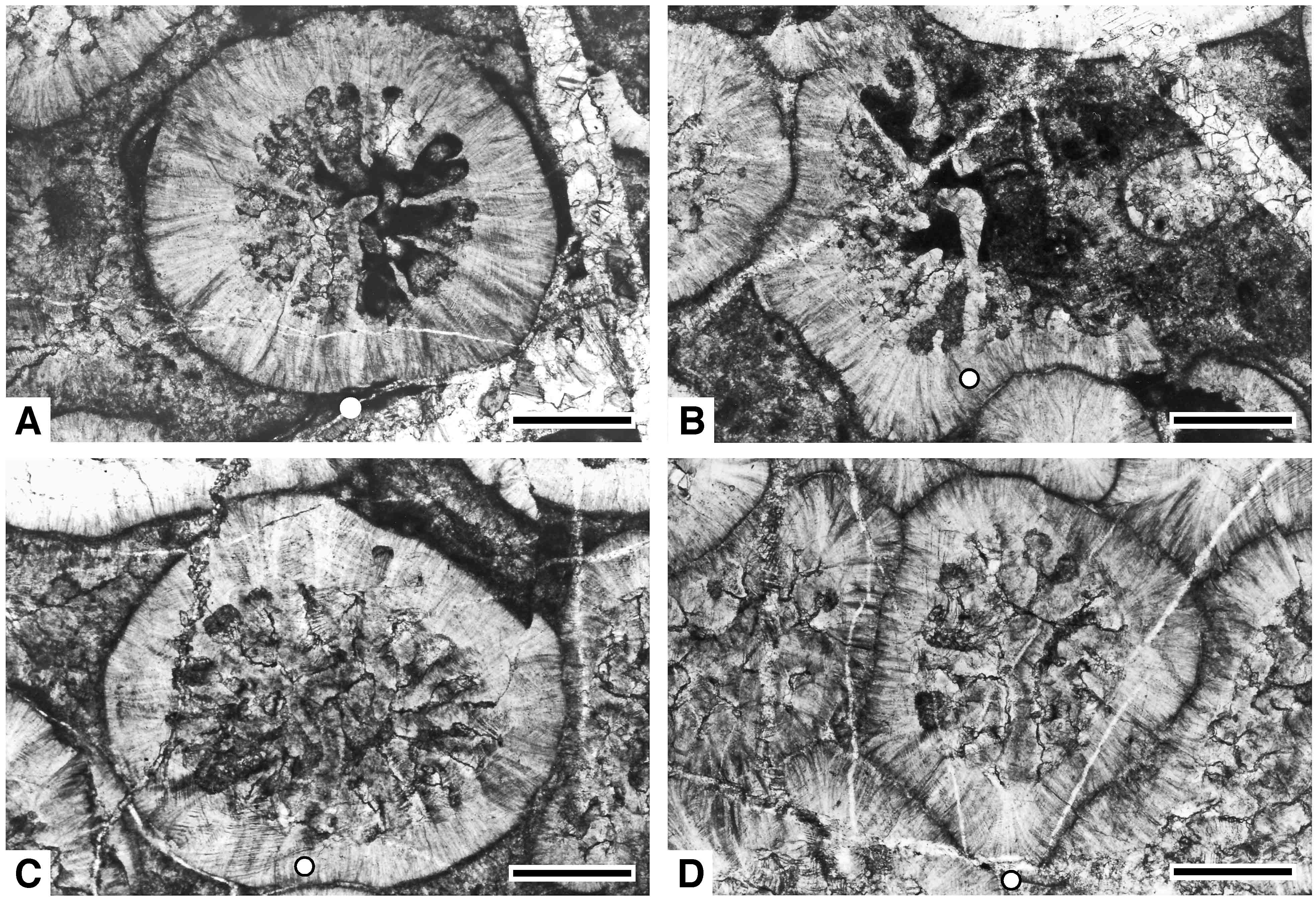
Yamatophyllum ultimumの壁の微細構造.繊維状の組織(初生)とジグザグ構造(後生)が顕著(Ezaki and Kato, 2014)
-3.jpg)
Rhizotrochus typusのrootletの形態的な特徴(Tokuda and Ezaki, 2013)
-4.jpg)
Rhizotrochus typusのrootletの微視的な特徴(Tokuda and Ezaki, 2013)
-9.jpg)
Truncatoflabellumのanthocyathus spineの巨視的から微視的な特徴(Tokuda and Ezaki, 2013)
生物間相互作用の地史的変遷
「骨格中に残された共生・寄生・競争・共創現象の解読」,「生物間関係性の変遷とその背後要因」,「棲息場の創出(開拓)過程の解析」


(左)現生六射サンゴ骨格に“寄生”するフジツボ.触手や繊毛が顕著.両者の関係は骨格中に記録される.(右)キッカサンゴがウミキノコをスウィーパー触手で攻撃している.


(左)同種の2群体の六射サンゴが隣接しながらも共存している.(右)同種の単体四射サンゴSchlotheimophyllumが隣接し,個体同士が癒合している.スウェーデン ゴトランド島 Visby (Wenlock)



単体Fungiaで見られる無性増殖後により形成された個体間の癒合.(左)見かけ上,縦分裂が生じたように見える. (中)口盤の裏側.柄部(anthocaulus)からの分離痕が2つ認められる.(右)口盤側で2個体の骨格間で癒合現象が認められる.



単体Fungiaで見られる無性増殖後により形成された個体間の癒合.(左)口盤側の軟体部.(中)口盤の裏側.柄部(anthocaulus)からの分離痕が2つ認められる.(右)口盤側で2個体の骨格間で癒合現象が認められる.


(左)ウニが底質の岩石を侵食している.(右)ウニが群体六射サンゴOulastreaを侵食している.和歌山県西牟婁郡白浜


(左)単体六射サンゴを穿孔する二枚貝(上部ジュラ系:和歌山県湯浅).(右)層孔虫と共成長する四射サンゴTryplasma. (上部シルル系Ludlow:スウェーデン ゴトランド島)


床版サンゴPleurodictyumの中にゴカイが共生している. (下部デボン系:ボリビア,Lapaz,Belen層)


巻貝の表面を被覆する群体六射サンゴSeptastrea marylandica.(左)上面.(右)同殻口側.(更新世Tamiami層:フロリダ州)


(左)単体六射サンゴHeterocyathus aequicostatus上面.(右)同サンゴ底面:孔の中に星虫が共生. (鹿児島県串本野沖100-150m)



(左)単体六射サンゴHeteropsammia cochlea上面.(中)同サンゴ底面:孔の中に星虫が共生.(右)同サンゴ側面.


(左)単体六射サンゴHeterocyathus japonicus上面.(右)同サンゴ側面:巻貝中の星虫と共生.(和歌山県日高郡南部沖120m)


(左)ヒドロサンゴJanaria mirabilis上面.(右)同サンゴ下面:孔の中にヤドカリ(Manucomplanus varians )が棲息している.スケール1cm.


(左)Conus magusの殻表面にOulastreaが付着(共成長).(右)Oulastreaの莢部の特徴.


(左)単体六射サンゴTruncatoflabellumの骨格表面での微生物類の穿孔痕.(右)同サンゴ骨格表面の付着生物(ゴカイ,コケムシ,単体サンゴなど):底質の二次的な活用 (熊本県天草沖水深30-50m).


サンゴ化石骨格(壁)中に微生物による穿孔痕が残されている.
(左)ペルム紀四射サンゴ:Ipciphyllum subelegans.マレーシア パハン 中部ペルム系(Kato and Ezaki, 1986).(右)古生代六射サンゴ:Houchangocyathus wangi.中国貴州省紫雲 中部ペルム系(Ezaki, 2000).
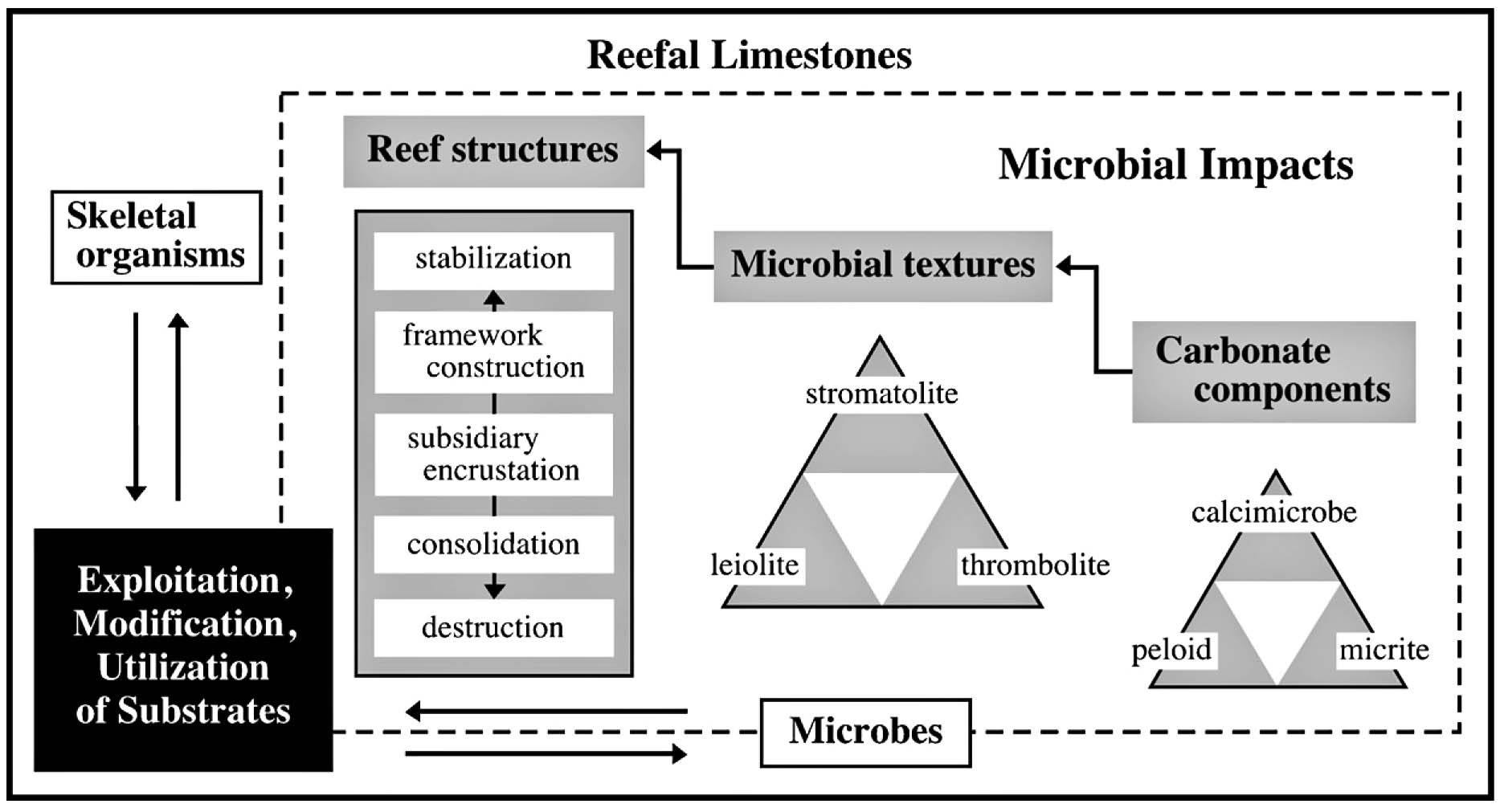
礁性石灰岩中の構成・構造・組織.微生物類と骨格生物との相互関係を考えながら,様々な観察レベル(スケール)で現象を捉える必要がある(Adachi and Ezaki, 2007).